
după cum puteți vedea în Figură1, nucleotidele variază doar ușor și numai în baza azotată. În cazul ADN – ului, aceste baze sunt adenina, guanina, citozina și timina. Observați similitudinea formelor de adenină și guanină, precum și similitudinea dintre citozină și timină. A și G sunt clasificate ca purine, în timp ce C și T sunt clasificate ca pirimidine. Atâta timp cât numim lucruri, observați „deoxiriboză” și „riboză”. După cum sugerează și numele, deoxiriboza este doar o riboză fără oxigen., Mai exact, în cazul în care există o grupare hidroxil atașată la carbonul 2 al ribozei, există doar un hidrogen atașat la carbonul 2 al dezoxiribozei. Aceasta este singura diferență între cele două zaharuri.în construirea aleatorie a unui singur fir de acid nucleic in vitro, nu există reguli particulare privind ordonarea nucleotidelor în raport cu bazele lor. Identitățile bazelor lor azotate sunt irelevante, deoarece nucleotidele sunt atașate prin legături fosfodiesterice prin gruparea fosfatică și pentoza., Prin urmare, este adesea menționată ca coloana vertebrală zahăr-fosfat. Dacă descompunem cuvântul „fosfodiester”, vedem că descrie destul de ușor conexiunea: zaharurile sunt conectate prin două legături esterice ( —O—) cu un fosfor între ele. Una dintre ideile care confundă adesea studenții este direcționalitatea acestei legături și, prin urmare, a acizilor nucleici în general. De exemplu, când vorbim despre ADN polimerază, enzima care catalizează adăugarea de nucleotide în celulele vii, spunem că funcționează într-o direcție 5-prime (5′) la 3-prime (3′)., Acest lucru poate părea ca arcane molecular-biolog-vorbesc, dar este de fapt foarte simplu. Uitați-vă la două dintre nucleotidele unite prin legătura fosfodiesterică (figura \(\PageIndex{1}\), stânga jos). O nucleotidă de adenină este îmbinată cu o nucleotidă de citozină. Legătura fosfodiesterică va lega întotdeauna 5-carbonul unei dezoxiriboze (sau riboză în ARN) cu 3-carbonul următorului zahăr. Acest lucru înseamnă, de asemenea, că la un capăt al unui lanț de nucleotide legate, va exista o grupare liberă de fosfat 5′ (- PO4), iar la celălalt capăt, o hidroxil 3′ liberă (-OH)., Acestea definesc direcționalitatea unui fir de ADN sau ARN.

ADN-ul este găsit în mod normal ca o moleculă de dublu-stranded în celulă întrucât ARN-ul este cea mai mare parte single-stranded., Este important să înțelegem totuși că, în condiții adecvate, ADN-ul ar putea fi făcut monocatenar, iar ARN-ul poate fi dublu-catenar. De fapt, moleculele sunt atât de asemănătoare încât este chiar posibil să se creeze molecule hibride dublu catenare cu un fir de ADN și unul de ARN. Interesant, ARN-ARN dublu helices și ARN-ADN dublu helices sunt de fapt puțin mai stabile decât cele mai convenționale ADN-ADN dublu helix.,baza naturii dublu catenare a ADN-ului și, de fapt, baza acizilor nucleici ca mediu pentru stocarea și transferul informațiilor genetice, este asocierea de bază. Împerecherea de bază se referă la formarea legăturilor de hidrogen între adenine și timine și între guanine și citozine. Aceste perechi sunt semnificativ mai stabile decât orice asociere formată cu celelalte baze posibile. Mai mult, atunci când aceste asociații de perechi de bază se formează în contextul a două fire de acizi nucleici, distanța lor este, de asemenea, uniformă și foarte stabilă., Vă puteți aminti că legăturile de hidrogen sunt legături relativ slabe. Cu toate acestea, în contextul ADN-ului, legarea hidrogenului este ceea ce face ca ADN-ul să fie extrem de stabil și, prin urmare, foarte potrivit ca mediu de stocare pe termen lung pentru informațiile genetice. Deoarece chiar și în procariote simple, helicele duble ale ADN-ului au cel puțin mii de nucleotide lungi, aceasta înseamnă că există câteva mii de legături de hidrogen care țin cele două fire împreună., Deși orice persoană nucleotide-să-nucleotide legături de hidrogen interacțiune ar putea fi ușor perturbat temporar de o ușoară creștere a temperaturii, sau o minuscula schimbare în forța ionică a soluției, un dublu-helix a ADN-ului necesită temperaturi foarte ridicate (în general peste 90oC) de a denatura complet dublu helix în componente individuale.deoarece există o împerechere exactă unu – la-unu a nucleotidelor, se dovedește că cele două fire sunt în esență copii de rezervă unul altuia-o plasă de siguranță în cazul în care nucleotidele sunt pierdute dintr-o singură șuviță., De fapt, chiar dacă părți ale ambelor fire sunt deteriorate, atâta timp cât celălalt fir este intact în zona de deteriorare, atunci informațiile esențiale sunt încă acolo în secvența complementară a firului opus și pot fi scrise în loc. Rețineți însă că, în timp ce un fir de ADN poate acționa astfel ca o „copie de rezervă” a celuilalt, cele două componente nu sunt identice – ele sunt complementare. O consecință interesantă a acestui sistem de fire complementare și antiparalel este că cele două fire pot purta fiecare informații unice.,perechile de gene Bi-direcționale sunt două gene pe fire opuse ale ADN-ului, dar care împărtășesc un promotor, care se află între ele. Deoarece ADN-ul poate fi făcut doar într-o singură direcție, 5′ la 3′, acest promotor bidirecțional, adesea o insulă CpG (vezi capitolul următor), trimite astfel ARN polimeraza pentru fiecare genă în direcții fizice opuse. Acest lucru a fost demonstrat pentru un număr de gene implicate în cancere (sân, ovarian) și este un mecanism de coordonare a expresiei rețelelor de produse genetice.catenele unui ADN dublu-helix sunt antiparalel., Aceasta înseamnă că, dacă ne uităm la o dublă helix de ADN de la stânga la dreapta, un fir ar fi construit în direcția 5′ la 3′, în timp ce firul complementar este construit în direcția 3′ la 5′. Acest lucru este important pentru funcția enzimelor care creează și repară ADN-ul, așa cum vom discuta în curând. În figura \(\PageIndex{1}\), partea stângă este 5 ‘la 3′ de sus în jos, iar cealaltă este 5’ la 3 ‘ de jos în sus.din punct de vedere fizic, moleculele de ADN sunt încărcate negativ (toți acești fosfați) și, în mod normal, o dublă helix cu o răsucire dreaptă., În această stare normală (numită și conformația „B”), o răsucire completă a moleculei cuprinde 11 perechi de baze, cu 0,34 nm între fiecare bază nucleotidică. Fiecare dintre bazele azotate sunt plane și, atunci când sunt asociate cu baza complementară, formează o „treaptă” plană at pe „scara” ADN-ului. Acestea sunt perpendiculare pe axa longitudinală a ADN-ului. Cea mai mare parte a ADN-ului plutitor liber într-o celulă și cea mai mare parte a ADN-ului în orice soluție apoasă de osmolaritate aproape fiziologică și pH, se găsește în această conformație B., Cu toate acestea, au fost găsite alte conformații, de obicei în circumstanțe de mediu foarte specifice. O conformație comprimată, A-ADN, a fost observată ca un artefact al cristalizării in vitro, cu puțin mai multe baze pe rând, o lungime mai scurtă și perechi de baze care nu sunt perpendiculare pe axa longitudinală. Un alt, Z-ADN, pare să se formeze tranzitoriu în întinderi de ADN bogate în GC, în care, interesant, ADN-ul se răsucește în direcția opusă.s-a sugerat că atât formele A cât și Z ale ADN-ului sunt, de fapt, relevante din punct de vedere fiziologic., Există dovezi care sugerează că forma A poate apărea în dublu helice hibride ARN-ADN, precum și atunci când ADN-ul este complexat la unele enzime. Conformația Z poate apărea ca răspuns la metilarea ADN-ului. Mai mult, conformația „normală” a ADN-ului B este o structură idealizată bazată pe faptul că este complet hidratată, așa cum este cu siguranță foarte probabil în interiorul unei celule. Cu toate acestea, această stare de hidratare se schimbă în mod constant, deși minuțios, astfel încât conformația ADN va varia adesea ușor de la parametrii de conformație B din Figura \(\PageIndex{2}\).,în procariote, ADN-ul se găsește în citoplasmă (destul de evident, deoarece nu există altă alegere în acele organisme simple), în timp ce în eucariote, ADN-ul se găsește în interiorul nucleului. În ciuda diferențelor dintre locațiile lor, nivelul de protecție împotriva forțelor externe și, mai ales, dimensiunile lor, atât ADN procariot cât și eucariot sunt ambalate cu proteine care ajută la organizarea și stabilizarea structurii cromozomiale globale., Relativ puțin se înțelege în ceea ce privește ambalajul cromozomial procariote, deși există asemănări structurale între unele dintre proteinele găsite în cromozomii procarioți și eucarioți. Prin urmare, cele mai multe cursuri introductive de biologie celulară se lipesc de ambalajul cromozomial eucariot.

ADN Gol, indiferent dacă sunt procariote sau eucariote, este un extrem de fir subțire de material, aproximativ 11 nm în diametru. Cu toate acestea, având în vedere dimensiunea genomului eucariot, dacă ADN-ul ar fi stocat în acest fel în interiorul nucleului, ar deveni încurcat. Imaginați-vă o găleată în care ați aruncat o sută de metri de fire, fără nici o încercare de a o organiza, înfășurându-l sau înfășurându-l., Acum, ia în considerare dacă v-ar fi capabil să ajungă în acea găleată trage pe un fir, și se așteaptă să trageți în sus doar un singur fir, sau dacă în schimb sunt susceptibile de a trage în sus cel puțin o mică încurcătură de fire. Celula face, în esență, ceea ce ați face cu firele pentru ao menține organizată: este ambalată perfect în jurubițe mai mici și mai ușor de gestionat. În cazul ADN-ului, fiecare cromozom este buclat în jurul unui complex histonic pentru a forma primul ordin de organizare cromozomială: nucleozomul.
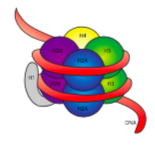
fibra de 30 nm este ținută împreună prin două seturi de interacțiuni. În primul rând, histonul linker, H1, aduce nucleozomii împreună într-o structură aproximativă de 30 nm., Această structură este apoi stabilizat prin legături disulfidice care se formează între histone H2A de unul nucleosome și H4 histone de vecinul său.histonele sunt o familie de proteine de bază (încărcate pozitiv). Toate acestea funcționează în principal în organizarea ADN – ului, iar nucleozomul se formează atunci când ADN-ul se înfășoară (puțin peste 2 ori) în jurul unui nucleu de opt histone-două fiecare dintre H2A, H2B, H3 și H4. Numărul și poziția de sarcini pozitive (cea mai mare parte din lysines și arginines) sunt cruciale pentru capacitatea lor de a lega strâns de ADN, care anterior subliniat, este foarte încărcat negativ., Că” contrariile atrag ” ideea nu este doar un sfat de dating din coloanele de sfaturi.

La o examinare de structura 3D a histone core complexe, vom vedea că în timp ce relativ neschimbate proteine interacțiune domenii țineți histones împreună în centru, încărcat pozitiv reziduuri sunt găsite în jurul afara de complexe, disponibile pentru a interacționa cu încărcat negativ fosfați de ADN.,
într-un capitol ulterior, vom discuta despre modul în care enzimele citesc ADN-ul pentru a transcrie informațiile sale pe bucăți mai mici, mai ușor de gestionat de ARN. Deocamdată, trebuie doar să fim conștienți de faptul că, la un moment dat, o mare parte din ADN este ambalată strâns, în timp ce unele părți ale ADN-ului nu sunt. Deoarece părțile disponibile pentru utilizare pot varia în funcție de ceea ce se întâmplă/în celulă la un moment dat, ambalajul ADN-ului trebuie să fie dinamic. Trebuie să existe un mecanism pentru a slăbi rapid legarea ADN-ului la histone atunci când ADN-ul este necesar pentru exprimarea genelor și pentru a strânge legarea atunci când nu este., După cum se dovedește, acest proces implică acetilarea și deacetilarea histonelor.

Histone Acetyltransferases (Pălării) sunt enzime care loc o acetil grup pe o lizină unei proteine histone. Grupările acetil sunt încărcate negativ, iar acetilarea nu numai că adaugă o grupare încărcată negativ, ci elimină și sarcina pozitivă din lizină. Acest lucru are ca efect nu numai neutralizarea unui punct de atracție între proteină și ADN, ci chiar respingerea ușoară a acestuia (cu sarcini similare)., Pe de altă parte a mecanismului, Histone Deactylases (HDACs) sunt enzime care elimina acetilare, și, astfel, a restabili interacțiunea dintre proteine histone și ADN-ul. Deoarece acestea sunt enzime atât de importante, este evident că nu li se permite să opereze vrând-nevrând pe orice histonă disponibilă și, de fapt, ele se găsesc adesea într-un complex cu alte proteine care controlează și coordonează activarea lor cu alte procese, cum ar fi activarea transcripției.