
Introducere
Telomerii sunt nucleoproteine structuri la marginile foarte liniar eucariote cromozomii. Ei rezolvă două probleme majore în același timp. Primul este despre protecția finală a cromozomilor. Se estimează că celulele umane normale trebuie să repare cel puțin 50 de pauze endogene dublu catenare (DSBs) pe celulă pe ciclu celular (Vilenchik și Knudson, 2003)., Telomerii distinge naturale cromozomiale capete de dăunătoare DSBs și pentru a preveni lor ectopice de reparații, de exemplu, de end-to-end fuziuni de cromozomi (vanSteensel și deLange, 1997). Al doilea este problema de replicare finală care se ocupă cu menținerea lungimilor telomere adecvate. Acest lucru a fost recunoscut independent de doi cercetători (Watson, 1972; Olovnikov, 1973). Deoarece polimerazele ADN replicative dependente de ADN nu pot finaliza sinteza ADN la capetele cromozomilor, compensarea pierderii secvenței telomerilor replicative trebuie să provină dintr-o ADN polimerază dependentă de ARN., Aceasta enzima, numita telomeraza, împreună cu primul telomerilor minisatellite secvență, a fost descoperit în ciliate Tetrahymena (Blackburn și Biliară, 1978; Greider și Blackburn, 1985). Cu toate acestea, acesta este doar un aspect al întreținerii lungimii telomerilor. Epigenetice regulamentul de lungimea telomerilor homeostaziei, inclusiv interacțiunea timp noncoding telomeric repeta conțin ARN și exonuclease activitate căi, de asemenea, au fost intens studiate datorită potențialului terapeutic (Wellinger et al., 1996; Polotnianka și colab., 1998; Pfeiffer și Lingner, 2012).,
telomeraza, enzima responsabilă de adăugarea secvențelor de repetare a telomerilor la capătul 3 ‘ al telomerilor, este o enzimă complexă conservată cu numeroase componente . În principiu, doar două componente principale sunt esențiale pentru activitatea enzimatică a telomerazei, o componentă proteică catalitică activă, numită telomere reverstranscriptază (terț) și o componentă șablon, formată din subunitatea telomerazei ARN (TR). În timp ce TERT este evolutiv destul de bine conservat, TR este foarte variabilă, cu lungimi variind de la ca. 150 nt (Tetrahymena) la mai mult de 2.000 nt (ciuperci din genul Neurospora)., Doar o regiune scurtă din întreaga moleculă TR servește ca șablon pentru ADN-ul telomerilor nou sintetizat (Greider și Blackburn, 1985; Qi et al., 2013). Această regiune din TR este de obicei formată dintr-un motiv complet de telomere urmat de unul parțial, acesta din urmă servind ca regiune de recoacere pentru ADN-ul telomerilor existent. Deși, în principiu, este necesară doar o singură nucleotidă suplimentară (ca motiv parțial), de obicei se găsesc mai multe. De exemplu, două nucleotide suplimentare formează motivul de recoacere la șoareci sau cinci la om (Blasco et al., 1995; Feng și colab., 1995)., Cu toate acestea, în plante, dimensiunea regiunii șablon este variabilă, de exemplu, două în Arabidopsis thaliana, șapte în Arabis sp. sau șase în Nicotiana (Fajkus et al., 2019). Celelalte regiuni TR au funcții structurale, de reglementare și proteine interactive . A se vedea, de asemenea, o reprezentare schematică a telomerazei și a ciclului său de activitate în Figura 1.

Figura 1 reprezentarea Schematică a telomerazei ciclu de activitate cu Arabidopsis-tip telomerilor șablon. TERT, Telomere reverstranscriptază; TR, subunitate de ARN telomerază., Figura bazată pe Sekhri (2014).
cât de variabile sunt secvențele telomerilor?
Telomerilor secvențe sunt de obicei scurte minisatellites tandemly aranjate, de obicei după formula (TxAyGz)n. La minisatellite aranjament provine din modul în care telomerazei sintetizează ADN-ul, pe scurt, și cea mai mare parte identice motive, unul câte unul., Mai multe ipoteze consideră că un astfel de aranjament este important deoarece promovează recunoașterea proteinelor specifice telomerilor de către homo – și heterodimeri și pentru potențialul de a forma cvadruplexuri G care pot stabiliza capetele cromozomilor sau pot servi ca substraturi pentru proteinele specifice telomerilor (Spiegel et al., 2020; Tran și colab., 2013). Secvențele de Telomere sunt bine conservate prin evoluție, iar grupuri mari de organisme folosesc motivul telomerilor tipic grupului pentru a-și construi ADN-ul telomerilor., Un număr tot mai mare de studii și proiecții mari au arătat că toate vertebratele testate și multe metazoane bazale folosesc TTAGGG (Meyne et al., 1989; Traut și colab., 2007), în timp ce Euarthropoda (artropode), inclusiv Hexapoda (insecte), au TTAGG (Frydrychova et al., 2004; Vitkova și colab., 2005). În mod constant, numeroase excepții se acumulează în timp, de exemplu, (A(G)1-8) în Dictyostelium (Emery și Weiner, 1981), TTAGGC în Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG în Coleoptera (gândaci) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGGG în Giardia (diplomonads) (Uzlikova et al.,, 2017), sau TTNNNNGGG în Yarrowia clade (drojdii) (Cervenak și colab., 2019). Mai mult, sistemele independente de telomerază, în care secvența telomerilor minisateliți a fost pierdută și înlocuită de repetări complexe, sunt reprezentate, de exemplu, de Diptera și Chironomidae (revizuite în (Mason et al., 2016)). Pentru o revizuire generală a secvenței telomere eucariote vezi (Fajkus et al., 2005; Fulneckova și colab., 2013).compoziția telomerilor în plante este și mai diversă., Aici vom folosi termenul de „plante” într-un sens larg, de asemenea, cunoscut sub numele de Archaeplastida sau regatul Plantae sensu lato, și cuprinde Rhodophyta (alge roșii), Glaucophyta, la Chlorophyte alge grad și Streptophyte alge clasa (cu totul cunoscut sub numele de alge verzi), și Embryophyta (plante terestre) (O Mie de Plante Transcriptomes Inițiativă, 2019). Secvența tipică de plante telomere este TTTAGGG, numită și tip Arabidopsis (sau pur și simplu, tip de plante), deoarece a fost descoperită în Arabidopsis thaliana (Richards și Ausubel, 1988) și acum în multe alte specii în aproape toate ordinele de plante., Deși TTTAGGG este încă cea mai frecventă, există o variabilitate semnificativă în secvențele telomerilor în liniile algelor roșii și verzi. Cum pentru alge roșii (Rhodophyta), telomerilor secvență de informații este cea mai mare lipsă sau fragmentar, deși unele telomerilor candidați au fost descoperite in silico, cum ar fi AATGGGGGG pentru Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T)AGGG pentru Galdieria sulphuraria (Fulneckova et al., 2013); TTAGGG a fost găsit în citirile genomice ale Porphyra umbilicalis (Fulneckova et al., 2013), dar sunt necesare mai multe dovezi pentru a confirma poziția lor terminală pe cromozomi., Diversitatea telomerilor în algele verzi reflectă atât schimbările dinamice, cât și caracterul său parafiletic. Deși TTTAGGG predomină în Chlorophyta, cum ar fi în genuri Ostreococcus (Derelle et al., 2006) și Chlorella (Higashiyama et al., 1995), multe alte motive divergente au fost detectate și acolo, cum ar fi TTAGGG în genul Dunaliella și Stephanosphaeria (Fulneckova et al., 2012), și TTTTAGGG în Chlamydomonas (Petracek și colab., 1990). În bazală Streptophyta (Klebsormidiophyceae) modificări progresive în motive de TTTAGGG să TTTTAGGG și TTTTAGG au fost descrise., Prezența TTAGGG în Rhodophyta și Glaucophyta duce la ipoteza că acest lucru este motiv de rudenie în plante (Archaeplastida) (Fulneckova et al., 2013).în ceea ce privește plantele terestre, una dintre primele proiecții efectuate a arătat că secvența de tip Arabidopsis a fost cea mai frecventă și a fost conservată în cea mai mare parte prin filogenia lor (Cox et al., 1993; Fuchs și colab., 1995), deși unii dintre acești autori au detectat deja mai multe excepții în familia Amaryllidaceae (fosta Alliaceae), în care secvența de tip Arabidopsis a fost absentă la mai multe specii., Mai târziu, prima telomerilor secvență neobișnuită pentru terenuri plante, vertebrate-tip TTAGGG, a fost caracterizat în Aloe și în alte Asparagales (Weiss și Scherthan, 2002; Puizina et al., 2003; Sykorova și colab., 2003c). A fost formulată o ipoteză despre pierderile și recuperările repetate ale secvenței telomerilor TTTAGGG și TTAGGG în Asparagales (Adams et al., 2001). Cu postrefinarea Ordinului Asparagales în APGIII (grupul de filogenie Angiosperm 2009) (Bremer et al.,, 2009), s-a arătat că numai două mari evolutiv switch-uri în telomerilor secvență de compoziție a avut loc (mai degrabă decât mai multe repetate pierderi și câștiguri), în următoarea ordine: primul din familia Iridaceae, în care trecerea de la planta-tip TTTAGGG la vertebrate de tip TTAGGG s-a întâmplat, urmată de familiile Xeronemataceae, Asphodelaceae și miezul Asparagales (inclusiv Amarillidaceae s.am și Asparagaceae s.l.,); și o a doua în subfamilia Allioideae (fostul tratat separat de familie, Alliaceae) în care un complet nou telomerilor secvență a apărut, CTCGGTTATGGG (Fajkus et al., 2016). În afara Asparagales, noi telomerilor secvențe au fost, de asemenea, detectat în teren a plantelor grupuri disparate ca (i) Solanaceae, în care telomerilor secvență de Cestrum elegans TTTTTTAGGG a fost descris (Sykorova et al., 2003a; Sykorova și colab., 2003b; Peska și colab., 2008; Peska și colab.,, 2015) și (ii) Lentibulariaceae, în cazul în care genul Genlisea a arătat o remarcabilă diversitate cu unele specii caracterizate prin Arabidopsis-tip telomerilor se repetă în timp ce altele expuse amestecate secvență variante TTCAGG și TTTCAGG (Tran et al., 2015).în ciuda tuturor excepțiilor de motive telomere detectate, diversitatea reală în secvențele telomerice din plantele terestre este probabil mult subestimată. O publicație recentă (Vitales et al.,, 2017), în care un screening de teren planta telomerilor secvențe fost efectuate, a constatat că telomerilor secvențe erau cunoscute doar de clar pentru mai puțin de 10% dintre specii și 40% din genurile conținute în Plantă adnr baza de date (www.plantrdnadatabase.com), o resursă furnizarea moleculară citogenetică informații despre plante de uscat (Garcia et al., 2012). Un rezumat al distribuției secvenței telomerilor în plante, după APG IV (grupul de filogenie Angiosperm, 2016) (Byng et al., 2016), precum și cea mai recentă filogenie vegetală (inițiativa O mie de Transcriptomi vegetali, 2019) se găsește în Figura 2.,
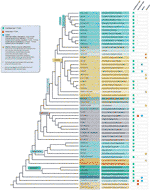
Figura 2 Telomerilor motive în Archaeplastida (plante în sens larg), bazat pe APG IV (Filogenia Grupului Angiospermelor 2016) și pe de O Mie de Plante Transcriptomes Inițiativă (2019). Lungimile ramurilor nu exprimă Scale în timp real., Pentru simplitate și pentru a economisi spațiu, anumite „grupuri” polifiletice (grade) marcate cu un asterisc în copac au fost reprezentate de o singură ramură; din același motiv, mai multe ordine minore (enumerate în pătratul albastru din partea superioară stângă a figurii) nu sunt reprezentate pe copac. Prima etichetă tip se referă de obicei la comenzi de plante și, în câteva cazuri, la diviziuni, grade și chiar familii; a doua etichetă afișează familii reprezentative și, în câteva cazuri, comenzi sau genuri reprezentative.
de la proiecții la descoperire: Cum pot fi identificate motivele Telomerice?,
dovada că o secvență de candidat dată este una telomerică reală include mai mulți pași care declară în mod corespunzător localizarea sa la toate terminalele cromozomiale și, eventual, implicarea telomerazei în sinteza sa. Citogenetica moleculară (mai ales prin hibridizare in situ fluorescentă, FISH) a devenit importantă pentru vizualizarea localizării terminale a sondelor marcate ale secvențelor candidate la toate terminalele cromozomiale. Cu toate acestea, pește independent nu este suficient pentru a dovedi poziția foarte terminal., De exemplu, AcepSAT356 a fost propus în onion ca candidat la telomere, pe baza rezultatelor analizei FISH (Pich și Schubert, 1998). Cu toate acestea, locația sa aparentă a terminalului de către in situ nu a fost niciodată legată în mod convingător de funcția telomere. De fapt, descoperirea de Allium minisatellite telomerilor secvență CTCGGTTATGGG și telomerazei ar însemna că AcepSAT356 este bandă terminală închisă, iar (Fajkus et al., 2019)., Pozitiv PEȘTE telomeric semnale pot masca, de asemenea, mici modificări în telomerilor motive, cum ar fi polimorfisme uninucleotidice sau rezultate fals negative pot duce la scurt telomerii fiind sub limita de detectie a tehnicii.există două abordări suplimentare care determină poziția terminală la o rezoluție mai mare decât peștele; acestea se bazează pe activitatea BAL31 a exonucleazei., Prima este cea clasică Terminal Fragmentelor de Restricție (TRF) analiza, în care probele tratate de BAL31 arăta scurtarea progresivă de terminal fragmente și o scădere a intensității semnalului cu cresterea timpului de exonuclease tratament. Analiza ulterioară a lungimilor fragmentelor se realizează prin hibridizarea Southern-blot (Fojtova et al., 2015). Al doilea este skimming-ul comparativ al genomului (date NGS) al ADN-ului genomic nedigerat și digerat BAL31, în paralel., În BAL31 tratat de date, există o semnificativă sub-reprezentare a telomerilor secvențe, prin urmare terminal secvențe sunt identificate prin comparație cu cele netratate set de date, folosind bioinformatica instrumente RepeatExplorer sau Tandem Finder .
celălalt test important al unui candidat dat de secvență de telomeri la o specie este demonstrarea activității telomerazei. În acest sens, o abordare experimentală utilă, dezvoltată mai întâi pentru celulele umane, este protocolul de amplificare a repetării telomerilor (TRAP) (Kim et al., 1994), urmată de secvențierea produselor detectate (Peska et al.,, 2015; Fajkus și colab., 2016), care este puțin mai puțin sensibil la rezultate fals pozitive decât peștele. Toate aceste metode, inclusiv pește (Fuchs et al., 1995; Shibata și Hizume, 2011) și altele, cum ar fi hibridizarea slot-blot (sykorova et al., 2003c) și TRAP (Fulneckova et al., 2012; Fulneckova și colab., 2016), poate fi folosit pentru a ecran pentru telomere în grupuri largi de organisme complexe, inclusiv plante., Cu toate acestea, numai o combinație de metode alese în mod corespunzător poate conduce în mod convingător la o concluzie cu privire la funcția de telomere a unei secvențe de candidat, deoarece rezultatele bazate pe o singură abordare ar putea fi înșelătoare. O imagine mai completă a strategiilor de novo telomerilor candidat secvența de identificare, inclusiv prima încercare în Tetrahymena (Greider și Blackburn, 1985) sunt rezumate într-un metodologice articol, cu accent pe NGS abordarea utilizată în centrale cu extrem de mare de genomuri (Peska et al., 2017).
există Homoplazie în secvențele telomerilor?,
secvența telomerilor ancestrali este considerată a fi TTAGGG și este cea mai frecvent întâlnită în arborele vieții (Fulneckova et al., 2013). Cu toate acestea, pare clar că frecvența homoplaziei în evoluția motivului telomerilor este relativ ridicată. De exemplu, scurte, simple motive ca planta de tip TTTAGGG au apărut în mod independent și în mod repetat în cryptomonads, oomycete ciuperci, și alveolates; în mod similar, vertebrate-tip TTAGGG a apărut în al doilea rând, în anumite grupe de plante (Asparagales, Rodophyta și Chlorophyta algele) (Sykorova et al., 2003c; Fulneckova și colab.,, 2012; Fulneckova și colab., 2013; Somanathan și Baysdorfer, 2018). Motivul pentru care unele telomerilor secvențe au apărut mai frecvent decât alte, de obicei, mai multe secvențe complexe este, probabil, legate de presiuni de selecție, care ar favoriza precizie pentru o anumită secvență specifică de ADN-proteine de interacțiune (Forstemann et al., 2003). Dacă a existat o schimbare în fiecare motiv telomer, interferența în structura nucleoproteinelor telomerice ar duce în mod necesar la instabilitatea genomului., Acesta este motivul telomerilor secvențe sunt atât evolutiv stabil, care cuprinde foarte puține roman și de succes secvențe, un model în concordanță cu ideea unor pierderi repetate și apariția tipic telomerilor secvențe, cum a fost propus pentru Asparagales (Adams et al., 2001).
descoperirea homoplaziei în secvențele telomerilor ridică întrebarea, care sunt cauzele și procesele moleculare care au loc în timpul acestor schimbări? O schimbare a secvenței telomerilor, în ciuda faptului că pare banală în unele cazuri (de ex.,, un T suplimentar), poate provoca interferențe grave cu integritatea genomului, din cauza unui echilibru perturbat în interacțiunile telomere ADN-proteină. De asemenea, nu este clar dacă o schimbare în secvența telomerilor poate avea vreun avantaj evolutiv; în acest sens, (Tran et al., 2015) a sugerat că apariția unei citozine „metilatabile” într-o componentă de telomere bogată în G ar ridica posibilitatea reglementării prin modificarea epigenetică.
care sunt motivele moleculare ale schimbărilor în motivele telomerilor?,
pentru a explica schimbarea secvenței telomerilor, primul candidat este subunitatea șablon a telomerazei, telomerazei ARN (TR). TR—ul identificat anterior din drojdie și vertebrate aparține unui grup diferit de transcrieri, a căror caracteristică de legătură a fost că au fost transcrise de ARN polimeraza II (Pol II) – în toate, cu excepția ciliaților; aceasta a fost singura excepție de la transcrierile Pol II înainte de publicarea identificării TR a plantei terestre . Folosind motivul telomerilor relativ lungi al Allium pentru a căuta TR-ul său în cadrul bazei de date a secvenței ARN totale, Fajkus și colab., (2019) a arătat că un caracterizat anterior noncoding ARN implicate în reacția de stres la A. thaliana, numit AtR8, a fost într-adevăr telomerazei ARN subunitate (Wu et al., 2012; Fajkus și colab., 2019). A fost o transcriere a ARN polimerazei III (Pol III) conținând elementele de reglementare corespunzătoare în structura sa promotoare. Multă vreme, cercetătorii s-au așteptat ca TR-ul plantelor să fie atât de divergent încât ar fi imposibil să-l identifice pe baza unei căutări omologice (Cifuentes-Rojas et al., 2011)., Cu toate acestea, un anumit grad de similitudine a fost utilizat cu succes pentru a identifica un TR comun în mai multe specii de Aliu cu explozie comparativă. În mod surprinzător, omologia de secvență, prezența acelorași elemente normative, precum și un șablon corespunzătoare regiune a condus la identificarea a TRs în Allium, Arabidopsis și mai mult de 70 de alte plante indepartate conexe, inclusiv a celor cu divergente telomerilor motive, cum ar fi Genlisea, Cestrum, și Tulbaghia., Din câte știm, nu există încă date despre nicio algă TR, ceea ce ar elucida dacă transcrierea Pol III A TR este o caracteristică generală pentru toate plantele sau nu. Această lucrare (Fajkus et al., 2019), bazat pe CRISPR knock-out și alte experimente, a arătat, de asemenea, că un candidat de ARN telomerază identificat anterior în A. thaliana (Cifuentes-Rojas et al., 2011; Beilstein și colab., 2012) nu a fost o subunitate șablon funcțional al telomerazei, așa cum a fost demonstrat, de asemenea, la scurt timp după (Dew-Budd et al., 2019)., Presupunând că dependența Pol II / Pol III pentru transcripția TR este un marker evolutiv fiabil, cercetările viitoare ale TR în alte linii eucariote principale vor deschide probabil noi perspective asupra originii eucariotelor. Telomeraza gene și telomerilor secvențe sunt nerecunoscute surse de informații în această direcție, și găsirea unui Pol III dependente TR biogeneza cale în ciliate și plante linii pot reprezenta primii pași în această direcție (Greider și Blackburn, 1989; Fajkus et al., 2019).
cum au devenit cromozomii liniari?,
marea majoritate a procarioților conțin cromozomi circulari, în timp ce cromozomii liniari sunt regula în eucariote., Prin urmare, există două scenarii posibile în care (i) linearizare a fost efectuată de către un primitiv telomerazei, care precede alte procese care au condus la curent liniar cromozomiale caracteristici și funcții sau (ii) linearizare de o pre-eucariote cromozom circular a fost inițial telomerazei independent, dar doar inainte de curent eucariote deviat, un primitiv telomerazei început să ocupe capetele cromozomilor și a devenit esențială pentru nou format liniar cromozomi (Nosek et al., 2006). Villasante și colab., (2007) au propus un scenariu evolutiv în care ruperea de rudenie procariote cromozom circular activat un mecanism de transpunere în ADN-ul se termină, permițând formarea telomerii de o recombinare dependente de replicare mecanism: consecințele acestei ipoteze a condus la concluzia surprinzatoare ca eucariote centromeres au fost derivate din telomerii.
Interesant, vizavi de procesul de liniarizare, de exemplu, formarea de cromozomi circulare (de asemenea, numit inel de cromozomi) a apărut din timp în timp în evoluția eucariote, deși este extrem de instabilă., De exemplu, în caz de Amaranthus tuberculatus, inel de cromozomi a apărut ca un stres indus de răspuns, care transportă rezistență împotriva unui erbicid (glifosat); plus inelul de cromozomi nu prezintă hibridizare cu telomerilor sonde în analiza cariotipului (Koo et al., 2018). Sistemul telomerazei aproape universal și excepționalitatea cromozomilor circulari din eucariote nu ne permit să susținem o ipoteză asupra celeilalte., Cu toate acestea, mașinile recombinaționale utilizate în prelungirea alternativă a telomerilor (ALT), o cale independentă de telomerază, asociată cu anumite tipuri de cancer uman (Zhang et al., 2019), este deja prezent în procariote. În plus, există dovezi de cromozom liniarizare care apar în mod independent în distinctă procariot linii (Ferdows și Barbour, 1989; Nosek et al., 1995; Volff și Altenbuchner, 2000). Prin urmare, ipoteza că primul cromozom eucariot liniar (originar dintr-un strămoș procariot) a fost independent de telomerază pare mai probabilă., Există câteva exemple care arată că telomerazei pe bază de sistem nu este esențială pentru menținerea telomerilor în toate eucariote: retrotransposons în Drosophila telomerii, tv satelit repetă în Chironomus, o altă insectă (Rubin, 1978; Biessmann și Mason, 2003), și în ALT telomerazei-negativ cancerele umane (Hu et al., 2016; Zhang și colab., 2019). Totuși, unele dintre aceste sisteme nu pot fi la fel de diferite, și poate au o origine comună: în Drosophila, telomerilor de întreținere, cu sediul în retrotransposition, nu este prea diferită de telomerazei pe bază de mecanism (Danilevskaya et al.,, 1998), ceea ce duce la ipoteza că telomeraza în sine poate fi un fost retrotransposon. Dar, cu siguranță, speciile de plante negative de telomerază nu au fost descoperite până în prezent și toate excepțiile, în care telomerul tipic de tip vegetal a fost absent, s-au dovedit mai târziu că au motive diferite, dar totuși sintetizate de telomerază. Cu toate acestea, utilajele ALT sunt prezente în plante în paralel cu activitatea telomerazei (Watson și Shippen, 2007; Ruckova et al., 2008). Întrebări interesante despre rolul telomerazei, telomerilor și întreținerea lor în tumorile plantelor apar din aceasta., Unul atractiv se referă la absența metastazelor la plante, în ciuda prezenței ALT, probabil legată de rigiditatea țesuturilor vegetale sau de sisteme imunitare diferite decât la animale (Seyfried și Huysentruyt, 2013).deși dobândim cunoștințe din ce în ce mai mari despre biologia telomerilor, încă nu putem explica apariția telomerazei în eucariote., Dovezile actuale susțin ipoteza că apariția eucariotelor împreună cu lor liniară cromozomi, telomerii, și telomeraza a fost legat de aspectul de spliceosomal introni în archaeal gazde (Koonin, 2006; Fajkus et al., 2019). Similitudinea dintre terț și alte retroelemente a fost discutată de ceva timp (Pardue et al., 1997). În mod remarcabil, un studiu relativ recent a arătat că TERT, ca membru probabil al grupului descendent ii introns, este secvențial aproape de retrotransposonii elementului Penelope (Gladyshev și Arkhipova, 2007)., Dar TERT este doar una dintre cele două componente esențiale ale telomerazei, iar TR este, la origine, și mai enigmatic datorită conservării secvenței sale scăzute în toate eucariotele .
concluzie
la începutul erei genomicii plantelor, secvența telomerilor a fost considerată aproape neschimbată. Conservarea generală a telomerilor și a sistemului de telomerază a sugerat că toate plantele pot avea telomere de tip tttaggg. Identificarea secvențelor neobișnuite de telomere în genomul complex al plantelor, în multe cazuri cu valori c gigantice (cum ar fi în Cestrum și Allium sp.,), a meritat efortul, deoarece motivul extrem de lung alium telomere a fost indiciul în căutarea unui TR autentic în plantele terestre. TR-ul nou descris în plante și cercetarea ulterioară a telomerilor/telomerazei în straturile bazale ale algelor ar putea dezvălui informații valoroase despre evoluția timpurie, prin urmare, cercetarea telomerilor din plante poate contribui semnificativ la ipoteze privind apariția eucariotelor.VP și SG au contribuit în mod egal la scrierea, editarea și pregătirea acestei mini-recenzii.,
finanțare
Conflict de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Aprecieri
O Mie de Plante Transcriptomes Inițiativă (2019). O mie de transcriptomi de plante și filogenomica plantelor verzi. Natura 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstract | CrossRef Textul Complet | Google Scholar