
Introducere
infecții Nosocomiale cauzate de tuberculoza multidrog rezistente Acinetobacter baumanni sunt din ce în ce mai frecvente în întreaga lume, în special în stabilirea de terapie intensiva (Wieland et al., 2018). Succesul acestei bacterii este facilitat de capacitatea sa de a supraviețui într-o varietate de medii agravate de capacitatea sa rapidă de a dobândi rezistență multidrog. Carbohidrații de suprafață joacă roluri cheie în starea generală de fitness și virulența A., baumannii (Lees-Miller și colab., 2013; Geisinger și Isberg, 2015; Weber și colab., 2016). A. baumannii produce polizaharidă capsulară cu greutate moleculară mare (CPS) care înconjoară membrana exterioară (Figura 1) (Russo et al., 2010). Cuprins de bine ambalate repetarea oligozaharide subunități (K unități), CPS forme discrete strat pe bacteriene de suprafață furnizarea de protecție de la diverse condiții de mediu, asistență în evaziune de gazdă de aparare imunitar și crește rezistența la o serie de compuși antimicrobieni (Russo et al., 2010; Iwashkiw și colab., 2012; Geisinger și Isberg, 2015).,
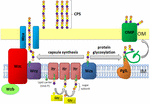
Figura 1. Reprezentarea schematică a ansamblului polizaharidic capsulă și exportul în A. baumannii. Sinteza începe cu transferaza inițială (Itr; maroon) situată în membrana interioară (IM) transferând primul zahăr al unității repetitive într-un purtător lipidic (Und-P; cerc verde). Zaharurile ulterioare sunt apoi adăugate la unitatea de creștere prin glicoziltransferaze specifice (GTR; galben) pe partea citosolică a membranei interioare., Subunitatea capsulei (unitatea K) este apoi transferată în periplasmă prin intermediul proteinei membranare integrale Wzx (Albastru închis). Subunitățile de zahăr sunt polimerizate de proteina wzy (violet) și complexul Wza/Wzb/Wzc (cyan, var, roșu) coordonează polimerizarea la nivel înalt și exportul lanțului de creștere, deplasându-le în membrana exterioară (OM). Pentru glicozilare, pgll (portocaliu) leagă unitățile K de proteinele membranei exterioare selectate (OMP; verde închis).în A. baumannii, asamblarea și exportul capsulelor are loc printr-o cale dependentă de Wzy (Hu et al.,, 2013; Kenyon și Hall, 2013; Willis și Whitfield, 2013; Woodward și Naismith, 2016) (Figura 1). De obicei, constând din zaharuri 4-6, unitatea K este asamblată pe molecula purtătoare de lipide undecaprenil pirofosfat (Und-P), care oferă o schelă pentru lanțul de zahăr în creștere (Whitfield, 2006). Primul zahăr din unitatea K este recrutat de o transferază inițială legată de membrană interioară (IM) (Itr), urmată de adăugarea secvențială de zaharuri la unitatea K în creștere de enzime specifice glicozil transferazei (GTR) (Figura 1) (Woodward și Naismith, 2016)., Fiecare K unitate este apoi transferat la periplasmic partea de IM de Wzx translocase și polimerizate prin Wzy, care transferă în creștere în lanț polizaharidic la o Und-P transportator la următoarea primite de subunitate (Figura 1) (Collins et al., 2007). După CPS polimer este sintetizat, este transportat la suprafata celulei printr-un extrem de coordonat proces care implică interacțiunea a trei proteine; Wza, Wzb, și Wzc, care cuprind mașini de export (Figura 1)., Sinteza CPS reprezintă un braț al unei căi bifurcate, deoarece aceste unități K sunt de asemenea utilizate pentru a decora anumite proteine de suprafață prin glicozilarea proteinei legate de o (Lees-Miller et al., 2013). În acest caz, o singură K unități sunt transferate la beneficiar proteine de O-oligosaccharyltransferase PglL (Figura 1) (Iwashkiw et al., 2012). În A. baumanni, glicozilarea proteinelor contribuie la formarea biofilmelor prin consolidarea inițială de atașament și de maturare de biofilme, și patogenitate cum s-a demonstrat într-un număr de animale infecție modele (Iwashkiw et al., 2012; Scott și colab., 2014; Harding și colab.,, 2015). Biofilmul este o stare de creștere în care comunitățile bacteriene sunt închise într-o matrice exopolizaharidică și sa dovedit a juca un rol semnificativ în persistența și rezistența A. baumannii.
Alte suprafața carbohidrati care sunt cunoscute că influențează patogenitatea A. baumanni includ lipooligosaccharide (LOS) și exopolysaccharide poli-β-(1-6)-N-acetilglucozamina (PNAG) (Preston et al., 1996; Weber și colab., 2016). PNAG formează „lipiciul” coeziv al biofilmelor și constituie o proporție substanțială de biofilme (Choi et al., 2009; Longo și colab., 2014)., Spre deosebire de majoritatea bacteriilor Gram-negative, A. baumannii nu produce lipopolizaharidă tradițională, ci în schimb un Glico-conjugat de suprafață similar, LOS, compus dintr-un miez lipidic A care nu are antigen O (Kenyon și Hall, 2013; Kenyon et al., 2014b). Pierderea producției LOS În A. baumannii scade stabilitatea membranei exterioare care duce la reducerea fitnessului (Moffatt et al., 2010; Beceiro et al., 2014).deși multe fragmente de carbohidrați influențează patogenitatea, se poate argumenta că CPS este un factor de virulență predominant al lui A. baumannii. Această revizuire își propune să consolideze ceea ce se știe despre A., capsula baumannii incluzând structurile selectate, biosinteza și organizarea genelor, rolul CPS în virulență și potențialul CPS ca țintă pentru dezvoltarea viitoare a vaccinului și a medicamentelor.pe măsură ce genomul complet al mai multor izolate A. baumannii devine disponibil, adevărata diversitate a structurilor capsulare prezente în această bacterie devine evidentă. Până în prezent, peste 100 de loci capsule unice (KL) au fost identificate în A. baumannii (figura 2a) (Shashkov et al., 2017). Aceste regiuni variază de obicei de la 20 la 35 kb în dimensiune., Analiza de gene regie CPS sinteza în zece completați A. baumanni genomul inițial a dus la denumirea de nouă tipurile de capsule, KL1–KL9, care a devenit baza pentru un universal tastarea schema pentru acești loci (Kenyon și Hall, 2013). Acest sistem s-a extins ulterior pentru a permite identificarea noilor loci K. Localizarea cromozomială a locusului K, între genele fkpa și lldP, este foarte conservată între tulpinile A. baumannii și conține acele gene necesare biosintezei și exportului specific fiecărui tip de CPS (Figura 2) (Hu et al.,, 2013; Kenyon și Hall, 2013). O excepție de la această regulă sunt A. baumanni tulpinile care posedă KL19 și KL39 regiuni, în cazul în care gena care codifică Wzy polimeraza, wzy, este găsit pe o mică genetice insulă în altă parte pe cromozom (Kenyon et al., 2016a). În plus, genele necesare pentru unele dintre zaharurile comune observate în CPS se găsesc în altă parte., Toate K loci arată un genetice similare de configurare, un foarte variabile grup de sinteză și transferaza genele necesare pentru biosinteza unic KL-tip zaharuri complexe, flancat pe o parte de înalt conservată CPS export gene, și pe de altă parte de un set de gene codifică conservate zaharuri simple și precursori (Figura 2A)., La wzx și wzy genele necesare pentru repetare-unitate de procesare sunt foarte variabile între K loci (Figura 2A, albastru deschis), care indică specificitatea pentru un anumit K unitate de structuri și, în general, ordinea de gtr determinanți, codare specifice glycosyltransferases cadrul KL regiuni, invers corespunde cu ordinea de acțiune.
regiunea variabilă a unor KL gene grupuri, cum ar fi KL37 și KL14, lipsa complex de gene pentru sinteza zaharurilor, deoarece acestea conțin doar zaharuri simple în K unități (Arbatsky et al., 2015; Kenyon și colab.,, 2015). De asemenea, adăugând la diversitatea lor, mai multe regiuni KL conțin gene redundante care nu sunt necesare pentru sinteza unității k finale. De exemplu KL8 și KL9 conține două itr gene (Kenyon și Hall, 2013) și KL37 are pgt1 phosphoglyceroltransferase, dar nu corespund phosphoglycerol reziduuri în a determinat structura (Arbatsky et al., 2015). În plus, în KL93, două de inserție secvență de elemente (ISAba26 și ISAba22) întrerupe pgt1 determinant (Figura 2) (Kasimova et al., 2017)., Genele necesare pentru biosinteza specifică a zahărului nu vor fi discutate, deoarece aceste căi au fost acoperite anterior și depășesc domeniul de aplicare al acestei revizuiri (Hu et al., 2013; Kenyon și Hall, 2013). În plus, alte gene situat în KL codifica produse estimată a fi implicate în acetilare sau acilare modificarea specifice glicanii (Figura 2A, roz). Deși examinarea loci K poate dezvălui multe despre structurile unității K, analiza chimică și testarea biochimică sunt necesare pentru a determina structurile exacte și pentru a identifica legăturile specifice dintre zaharurile repetate.,primele studii privind serotiparea CPS a lui A. baumannii au fost determinate de necesitatea dezvoltării unei metode de discriminare a izolatelor A. baumannii de alte specii de Acinetobacter, deoarece analiza fenotipică a fost împovărată de ambiguitate și identificare greșită (Traub, 1989). Așa cum s-a descris mai sus, există o diversitate fenomenală observată în grupurile de gene de biosinteză A. baumannii CPS, ceea ce se traduce în diversitatea observată în structura unității K (Hu et al., 2013; Kenyon și Hall, 2013). Colectiv, peste 40 diverse A., structurile unității baumannii K au fost elucidate până acum folosind spectroscopia RMN și analiza chimică. Structurile unitare K diferă în compoziția zahărului. Acestea pot include derivați ai zaharurilor comune legate de UDP, cum ar fi glucoza, galactoza și acidul glucuronic sau zaharuri rare și atipice, cum ar fi acizii non-2-ulosonici. Structurile variază în lungime și pot consta doar din două reziduuri, așa cum se vede pentru K53 tip CPS (Shashkov et al., 2018), sau până la cinci sau șase monozaharide, cum ar fi cea observată în K37 (figura 2b) (Arbatsky et al., 2015)., Structurile diferă, de asemenea, în legăturile atât în interiorul, cât și între unitățile K, rezultând producția de unități K care sunt liniare sau implică ramuri laterale, așa cum se vede în K1 și, respectiv, K93 (figura 2b) (Kenyon et al., 2016a; Kasimova și colab., 2017). Diferențele în localizarea legăturilor glicozidice specifice și a modelelor de o-acetilare ale diferitelor oligozaharide în cadrul unei structuri contribuie, de asemenea, la diversitatea unității K.,
Variație între K unitate de structuri pot fi subtile, de exemplu, K12 și K13 diferă numai prin comasarea a două glicanii, care necesită utilizarea de un supleant Wzy polimeraza; în consecință, K loci de ambele tulpini sunt identice, cu excepția pentru wzy gene (Figura 2). Alternativ, variația poate fi izbitoare, cum ar fi includerea de rare zaharuri, inclusiv pseudaminic, legionaminic, sau acinetaminic derivați de acid cum se vede în K2/6, K49, și K12/13 structuri, respectiv (Figura 2) (Kenyon et al., 2014a, 2015; Vinogradov și colab., 2014; Kasimova și colab., 2018)., Interesant este că derivații acidului acinetaminic au fost identificați doar în A. baumannii și nu se găsesc nicăieri altundeva în natură (Kenyon et al., 2017). În plus, unele K unități includă unic derivate specifice glicanii, de exemplu pseudaminic acid de K93 este acetilat cu o (R)-3-hydroxybutanoyl grup (Kasimova et al., 2017), în timp ce în k2 și K6 acidul pseudaminic este non-acetilat (Figura 2)., În plus, structura de K4 este unic, deoarece conține doar aminosugars, D-QuipNAc, și un terminal de N-acetil-D-galactozamină (D-GalpNAcA) ramură care este acoperit cu un pyruvyl grup, de o rară motiv și prima care urmează să fie descrise în Acinetobacter (Figura 2A, negru) (Kenyon et al., 2016b). Pe măsură ce numărul structurilor unitare K crește elucidat, la fel și încrederea de a deduce structura unitară K din analiza clusterelor de biosinteză poziționate în A. baumannii KL., Cu toate acestea, deși informativ, o înțelegere a rolului CPS joacă în patogeneza este important să se aplice aceste cunoștințe pentru îmbunătățirea rezultatelor infecțiilor A. baumannii.
rol în virulență, rezistență antimicrobiană și persistență
este fără îndoială că prezența CPS este esențială pentru patogenitatea A. baumannii. Nu numai că este necesar pentru evaziunea de apărare imună gazdă (Russo et al., 2010; Geisinger și Isberg, 2015), dar este important pentru rezistența la compuși antimicrobieni și supraviețuirea în medii adverse (Luke et al., 2010; Russo și colab.,, 2010; Geisinger și Isberg, 2015). CPS mediază evaziunea imună în multe tulpini A. baumannii prin limitarea interacțiunilor dintre structurile imunogene de suprafață ale bacteriilor și apărarea gazdei (Preston et al., 1996; Wu și colab., 2009; Russo și colab., 2010; Umland și colab., 2012; Lees-Miller și colab., 2013; Geisinger și Isberg, 2015; Wang-Lin și colab., 2017). Eliminarea capsulei în mai multe tulpini diferite de A. baumannii a arătat o supraviețuire redusă în serul uman și lichidul de ascită și atenuarea modelelor de infecție la șobolan și murină (Russo et al., 2010; Umland și colab., 2012; Lees-Miller și colab.,, 2013; Sanchez-Larrayoz și colab., 2017). În plus, până-regulamentul de capsule de producție în general utilizat A. baumanni tulpina ATCC 17978 (K3 CPS tip) serice crescute de rezistență și virulență într-un mouse-ul infecție model (Geisinger și Isberg, 2015). Mai mult decât atât, romanul tratamente antimicrobiene ar putea fi dezvoltate pentru anumite CPS tipuri, de exemplu cele care conțin pseudaminic acid, ca prezența sa a fost corelată cu virulență sporită (Hitchen et al., 2010; Kao și colab., 2016).
pe lângă protecția împotriva apărării gazdă, în A., producția baumannii CPS crește rezistența la o serie de compuși antimicrobieni, inclusiv cei utilizați pentru dezinfecție în setările clinice (Geisinger și Isberg, 2015; Tipton et al., 2015; Chen și colab., 2017). În plus, creșterea A. baumannii în nivelurile sub-inhibitorii de antimicrobiene influențează producția de CPS. De exemplu, expunerea la antibiotice-cloramfenicol sau eritromicină condus la îmbunătățirea capsule de sinteză în ATCC 17978 (Geisinger și Isberg, 2015) și meropenem de expunere selectat pentru mutații care duc la o pierdere în CPS de producție în izola 37662 (Chen et al., 2017)., Studiile efectuate pe o gamă mai largă de tulpini de A. baumannii sunt necesare pentru a identifica dacă protecția împotriva antimicrobienelor oferită de CPS este specifică tulpinii, specifică tipului de capsulă sau universală.capacitatea lui A. baumannii de a persista în mediul clinic a sporit, fără îndoială, colonizarea și infecțiile la pacienții susceptibili. A. baumannii este capabil să supraviețuiască luni întregi pe suprafețe spitalicești, cum ar fi șine de pat, mobilier și dispozitive medicale, oferind un rezervor care este adesea sursa de transmitere și infecție (Wendt et al., 1997; Gayoso și colab., 2013)., Toleranța remarcabilă de desicare a lui A. baumannii se crede că se datorează unei strategii de „bust sau boom”, în care o subpopulație persistentă de celule supraviețuiește în detrimentul celulelor moarte; CPS îmbunătățește toleranța la desicare prin furnizarea unei bariere fizice care facilitează retenția apei (Roberts, 1996; Webster et al., 2000; Gayoso și colab., 2013; Bravo și colab., 2016). Un rol direct al CPS în rezistența la deshidratare a fost demonstrat recent în tulpina A. baumannii AB5075 (Tip K25 CPS). În acest studiu, varianta acapsulară a AB5075 a afișat un 2.,Reducerea de 5 ori a viabilității în comparație cu tulpina parentală (Tipton et al., 2018). În plus, în două rude apropiate ale A. baumanni, Acinetobacter calcoaceticus și Acinetobacter baylyi, producția de exopolysaccharide și/sau CPS a fost demonstrat de a promova deshidratarea supraviețuire (Roberson și Firestone, 1992; Ophir și Gutnick, 1994). În plus față de influențarea rezistenței la deshidratare, CPS a fost asociat cu alte trăsături de virulență, inclusiv motilitatea (McQueary et al ., 2012; Huang și colab., 2014) și producția de biofilm (Umland et al., 2012; Lees-Miller și colab.,, 2013), cimentându-și astfel rolul de factor patogen.
studii Recente au legat de fază variabilă fenotip de A. baumanni AB5075 cu modificări în CPS de producție, ca extrem de virulent opac variante produce o CPS strat cu de două ori grosimea lor translucid omologii (Chin et al., 2018). Această tranziție de la translucid la opac a crescut, de asemenea, dramatic potențialul patogen al lui A. baumannii AB5075. Rezistența la dezinfectanții spitalicești obișnuiți și un subset de antibiotice aminoglicozide au fost, de asemenea, crescute (Tipton et al., 2015; Chin și colab.,, 2018) și opac variante au fost, de asemenea, mai rezistent la om lizozim, a cathelicidin legate de peptida antimicrobiană LL37 și peroxid de hidrogen, comparativ cu colonii translucide (Chin et al., 2018). Mai mult, izolatele opace au avut o toleranță crescută la condițiile deshidratate și la omologii translucizi în afara concurenței într-un model de infecție cu șoarece (Chin et al., 2018). Deoarece sunt implicați mai mulți factori în variația de fază, au fost efectuate studii suplimentare pentru a determina contribuția producției de CPS la fenotipul opac mai virulent., În urma publicării, autorii au demonstrat că o acapsular variantă a fost semnificativ mai sensibile la lizozim și dezinfectanți în comparație cu opac AB5075 de tip sălbatic părinte (Tipton et al., 2018). Interesant, nu a fost nicio diferență în rezistența la LL-37 și peroxid de hidrogen între opace de tip sălbatic și acapsular tulpini, sugerând alți factori decât CPS de producție duce la acest fenotip opac pentru variante de A. baumanni (Tipton et al., 2018).,
reglarea producției de CPS
indicii de mediu, cum ar fi temperatura, presiunea osmotică și modificările disponibilității metabolitului și ionilor pot influența producția de CPS bacterian (Hagiwara et al., 2003; Lai și colab., 2003; Mouslim și colab., 2004; Willenborg et al., 2011). Este surprinzător faptul că de câteva mecanisme de reglementare au fost identificate pentru CPS de producție ca CPS niveluri sunt de obicei reglementate post-translațional prin fosforilarea CPS export utilaje (Whitfield și Paiment, 2003; Chiang et al., 2017). În A., baumanni, doar două regulatoare de capsule de producție au fost până acum identificate; BfmRS și OmpR-EnvZ, ambele două componente de transducție a semnalului sisteme care joacă roluri multiple de reglementare implicate în plic biogeneza (Geisinger și Isberg, 2015; Tipton și, mai Degrabă, 2017; Geisinger et al., 2018). Atunci când este supus la antibiotice presiune, A. baumanni ATCC 17978 cps exprimare a fost crescut într-o BfmRS manieră dependentă (Geisinger și Isberg, 2015). Variația de fază, și, astfel, potențial de producție CPS, este puternic reglementată de sistemul OmpR-EnvZ în A., baumanni AB5075, ca mutații în oricare OmpR sau EnvZ a dus la o creștere semnificativă opac la translucid cu o frecvență de comutare (Tipton și, mai Degrabă, 2017). Deși tranziția de la translucid la opac are ca rezultat o creștere de două ori a grosimii capsulei, analizele transcriptomice nu au identificat diferențe în nivelurile de Expresie ale genelor KL între cele două faze.
CPS ca țintă pentru dezvoltarea vaccinurilor și tratamentelor împotriva A. baumannii
Antibiotic (în special cabapenem) rezistent A., baumannii sunt clasificate ca fiind un organism critic prioritar 1 al Organizației Mondiale a Sănătății pentru dezvoltarea de noi antimicrobiene (OMS, 2017). Deși nu există tratamente non-antibiotice sau vaccinuri autorizate pentru A. baumannii în prezent, există un interes crescut pentru dezvoltarea lor, iar studiile preliminare arată promițătoare. Expunerea la suprafață și prevalența tulpinilor patogene de A. baumannii fac din CPS o țintă ideală atât pentru tratamentele antimicrobiene, cât și pentru vaccinuri (Russo et al., 2013)., Acestea includ dezvoltarea de terapii pe bază de anticorpi, cum ar fi vaccinurile profilactice, imunizarea pasivă și terapia cu fagi (García-Quintanilla et al., 2013).mai multe studii au arătat eficacitatea imunizării pasive la șoareci folosind un anticorp specific CPS, care este protector împotriva provocării bacteriene cu 13-55% din izolatele clinice A. baumannii (Russo et al., 2013; Yang și colab., 2017; Lee și colab., 2018)., În plus, inocularea cu vaccinuri conjugate care încorporează glicani CPS atașați la un purtător de proteine obține o protecție imună mai bună decât CPS purificat împotriva unei game diverse de tulpini A. baumannii (Yang et al., 2017).interesul pentru terapia cu fagi pentru tratarea infecțiilor bacteriene a crescut în ultimii ani ca răspuns la criza actuală de creștere a rezistenței antimicrobiene. Terapia cu fagi este atractivă ca o potențială cale de tratament pentru infecțiile cu A. baumannii rezistente la mai multe medicamente., De exemplu, s-a constatat că un fag care codifică o depolimerază CPS degradează CPS de aproximativ 10%, patru din 38, rezistente la medicamente clinice A. baumannii testate (Hernandez-Morales et al., 2018). Deși gama gazdă a acestui fag este limitată, ar putea fi încorporată ca parte a unui cocktail de fag pentru a maximiza eficacitatea sau a viza focare specifice (Hernandez-Morales et al., 2018). Un fag care scindează selectiv legătura lui A., baumannii CPS la o ramură de acid pseudaminic poate fi valoroasă pentru terapia cu fagi sau pentru fabricarea eficientă a CPS purificat pentru dezvoltarea vaccinului și a anticorpilor (Lee et al., 2018). Fagul care vizează A. baumannii s-a dovedit recent a fi stabil atunci când este impregnat în produsele de îngrijire a rănilor arse într-o serie de condiții, inclusiv în prezența antimicrobienelor (Merabishvili et al., 2017). Cu toate acestea, deoarece majoritatea studiilor care investighează A. baumannii CPS sunt de obicei limitate la o anumită tulpină A. baumannii, nu se știe dacă aceste constatări se traduc la toate A., baumannii izolate sau dacă acestea sunt tulpina sau tipul capsulei specifice. Înțelegerea rolurilor pe care le joacă capsula în mai multe tulpini este esențială în identificarea tipurilor CPS care reprezintă cele mai bune ținte pentru noile vaccinuri sau dacă dezvoltarea antimicrobienelor care vizează calea biosintezei capsulelor este într-adevăr chiar fezabilă.deși capsula reprezintă o trăsătură importantă de virulență a lui A. baumannii, există date limitate disponibile privind rolul diferitelor tipuri de CPS în provocarea bolii., Pentru a dezvolta vaccinuri și terapii eficiente care vizează CPS, trebuie mai întâi să obținem o înțelegere cuprinzătoare față de mecanismele din spatele sintezei și expresiei sale, alături de avantajele pe care capsula le transmite bacteriilor gazdă. Această cercetare trebuie abordată în contextul variației extreme a serotipurilor CPS găsite în A. baumannii, pentru a asigura funcționarea potențialelor intervenții împotriva tulpinilor care produc diverse structuri CPS., Sunt necesare studii suplimentare privind CPS pentru a oferi o platformă pentru dezvoltarea de măsuri preventive și tratamente împotriva acestui agent patogen uman din ce în ce mai persistent și mortal.
contribuțiile autorului
JS a scris primul proiect. MB furnizat de intrare academice și revizuirea critică a articolului. FA produs aliniamente genomului și cu condiția revizuirea critică a manuscrisului. Toți autorii au aprobat versiunea finală.
finanțare
această lucrare a fost susținută de un grant Flinders Medical Research Foundation pentru MB. FA a fost susținută de AJ și IM Naylon și Satisface Încredere Doctorat., Burse. JS a fost susținută de o bursă AJ și IM Naylon.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Traub, W. H. (1989). Serotiparea Acinetobacter baumannii pentru delimitarea focarelor de infecție încrucișată nosocomială. J. Clin. Microbiol. 27, 2713–2716.
PubMed Abstract | Google Scholar
Whitfield, C., și Paiment, A. (2003)., Biosinteza și asamblarea polizaharidelor capsulare din grupa 1 în Escherichia coli și a polizaharidelor extracelulare asociate în alte bacterii. Ne. Rez.338, 2491-2502. doi: 10.1016 / j. carres.2003.08.010
PubMed Abstract | CrossRef Textul Complet | Google Scholar
CINE (2017). Lista prioritară globală a bacteriilor rezistente la antibiotice pentru a ghida cercetarea, descoperirea și dezvoltarea de noi antibiotice. Geneva: Organizația Mondială A Sănătății.
Google Scholar