
jak můžete vidět na Obrázku1, nukleotidy se liší jen mírně, a to pouze v dusíkaté báze. V případě DNA jsou tyto báze adenin, guanin, cytosin a thymin. Všimněte si podobnosti tvarů adenin a guaninu a také podobnosti mezi cytosinem a thyminem. A A G jsou klasifikovány jako puriny, zatímco C A T jsou klasifikovány jako pyrimidiny. Dokud pojmenováváme věci, všimněte si „deoxyribózy“a “ ribózy“. Jak název napovídá, deoxyribóza je jen ribóza bez kyslíku., Konkrétněji, kde je hydroxylová skupina připojená k 2-uhlíku ribózy, je k 2-uhlíku deoxyribózy připojen pouze vodík. To je jediný rozdíl mezi těmito dvěma cukry.
V náhodně konstrukci jednovláknové nukleové kyseliny in vitro, tam jsou žádná zvláštní pravidla týkající se pořadí nukleotidů s ohledem na své základny. Identita jejich dusíkatých bází je irelevantní, protože nukleotidy jsou připojeny fosfodiesterovými vazbami přes fosfátovou skupinu a pentózu., Proto se často označuje jako kostra cukru a fosfátu. Pokud rozebereme slovo „fosfodiester“, vidíme, že docela dobře popisuje spojení: cukry jsou spojeny dvěma esterovými vazbami ( —O—) s fosforem mezi nimi. Jednou z myšlenek, která často zaměňuje studenty, je směrovost této vazby, a tedy nukleových kyselin obecně. Například, když mluvíme o DNA polymeráze, enzymu, který katalyzuje přidání nukleotidů do živých buněk, říkáme, že funguje ve směru 5-prime (5′) až 3-prime (3′)., To se může zdát jako tajemný molekulární biolog-mluvit, ale ve skutečnosti je to velmi jednoduché. Další pohled na dva nukleotidy Spojené fosfodiesterovou vazbou (obrázek \(\PageIndex{1}\), vlevo dole). Adenin nukleotid je spojen s cytosinovým nukleotidem. Fosfodiesterová vazba vždy spojí 5-uhlík jedné deoxyribózy (nebo ribózy v RNA) s 3-uhlíkem dalšího cukru. To také znamená, že na jednom konci řetězce propojených nukleotidů bude volná 5′ fosfátová (-PO4) skupina a na druhém konci Volný 3′ hydroxyl (-OH)., Ty definují směrovost řetězce DNA nebo RNA.

DNA se obvykle nachází jako dvouvláknová molekula v buňce, zatímco RNA je většinou jednovláknová., Je však důležité pochopit, že za vhodných podmínek by mohla být DNA vytvořena jednovláknová a RNA může být dvouvláknová. Ve skutečnosti jsou molekuly tak podobné, že je dokonce možné vytvořit dvouvláknové hybridní molekuly s jedním pramenem DNA a jedním z RNA. Zajímavé je, že dvojité spirály RNA-RNA a dvojité spirály RNA-DNA jsou ve skutečnosti o něco stabilnější než konvenčnější dvojitá šroubovice DNA-DNA.,
Na základě dvouvláknové povaze DNA, a ve skutečnosti na základě nukleových kyselin jako médium pro ukládání a přenos genetické informace, je párování bází. Báze-párování se týká tvorby vodíkových vazeb mezi adeniny a thyminy a mezi guaniny a cytosiny. Tyto páry jsou výrazně stabilnější než jakákoli asociace vytvořená s ostatními možnými základnami. Kromě toho, když tyto base-pair sdružení podobě v rámci dvou vláken nukleových kyselin, jejich rozestup je také jednotné a vysoce stabilní., Možná si vzpomenete, že vodíkové vazby jsou relativně slabé vazby. V souvislosti s DNA je však vodíková vazba tím, co činí DNA extrémně stabilní, a proto je vhodná jako dlouhodobé úložné médium pro genetickou informaci. Protože i v jednoduchých prokaryot, DNA dvojité šroubovice jsou přinejmenším tisíce nukleotidů dlouhý, to znamená, že existuje několik tisíc vodíkových vazeb drží dva kusy dohromady., I když každý jednotlivý nukleotid-nukleotid vodíkové vazby interakce by mohla snadno být dočasně narušen mírným zvýšením teploty, nebo nepatrné změny v iontové síle roztoku, plný double-helix DNA vyžaduje velmi vysoké teploty (obvykle nad 90oC) zcela denaturaci dvoušroubovice na jednotlivá vlákna.
Protože tam je přesný one-to-one párování nukleotidů, ukázalo se, že dvě vlákna jsou v podstatě záložní kopie sebe – bezpečnostní síť v případě, že nukleotidy jsou ztraceny ze jeden pramen., Ve skutečnosti, i když jsou části obou pramenů poškozeny, pokud je druhý pramen neporušený v oblasti poškození, pak jsou základní informace stále v komplementární sekvenci opačného pramene a mohou být zapsány na místo. Mějte však na paměti, že zatímco jeden řetězec DNA může působit jako“ záloha “ druhého, oba prameny nejsou totožné – doplňují se. Zajímavým důsledkem tohoto systému komplementárních a antiparalelních pramenů je to, že oba prameny mohou nést jedinečné informace.,
obousměrné genové páry jsou dva geny na opačných pramenech DNA, ale sdílejí promotor, který leží mezi nimi. Od DNA mohou být provedeny pouze v jednom směru 5′ k 3′, tento bi-directional promotor, často CpG ostrova (viz další kapitola), tak odešle RNA polymeráza pro každého genu v opačné fyzikální směry. To bylo prokázáno u řady genů zapojených do rakoviny (prsu, vaječníků) a je mechanismem pro koordinaci exprese sítí genových produktů.
prameny dvojité šroubovice DNA jsou antiparalelní., To znamená, že pokud jsme se podívali na double-helix DNA zleva doprava, jeden pramen by mohl být postaven v 5′ 3′ směr, zatímco komplementární pramen je zhotoven v 3′ k 5′ směr. To je důležité pro funkci enzymů, které vytvářejí a opravují DNA, jak budeme brzy diskutovat. Na obrázku \(\PageIndex{1}\) je levý pramen 5′ až 3′ shora dolů a druhý je 5′ až 3′ zdola nahoru.
Z fyzikálního hlediska, molekuly DNA jsou záporně nabité (všechny ty fosfáty), a obvykle double-helix s pravák twist., V tomto normální (také volal „B“ konformace) stát, jeden vrut molekuly zahrnuje 11 párů bází, s 0,34 nm mezi každým nukleotidů základny. Každá z dusíkatých bází je rovinná a při spárování s komplementární bází tvoří na „žebříku“ DNA rovinnou „příčku“. Ty jsou kolmé k podélné ose DNA. Většina free-plovoucí DNA v buňce, a většina DNA v každém vodném roztoku téměř fyziologické osmolarity a pH, je nalezen v B konformaci., Byly však nalezeny další konformace, obvykle za velmi specifických environmentálních okolností. Jako artefakt krystalizace in vitro byla pozorována komprimovaná konformace, a-DNA, s o něco více bází na otáčku, kratší délka otočení a páry bází, které nejsou kolmé k podélné ose. Další, Z-DNA, se zdá, že se přechodně tvoří v úsecích DNA bohatých na GC, ve kterých se zajímavě DNA otáčí opačným směrem.
bylo navrženo, že jak a, tak z formy DNA jsou ve skutečnosti fyziologicky relevantní., Existují důkazy, které naznačují, že forma může dojít v RNA-DNA hybridu dvojité šroubovice, stejně jako když je DNA komplexu do některé enzymy. Z konformace může nastat v reakci na methylaci DNA. Kromě toho je „normální“ konformace B-DNA něco jako idealizovaná struktura založená na úplné hydrataci, což je jistě velmi pravděpodobné uvnitř buňky. Nicméně, hydrataci, stav se neustále mění, byť jen nepatrně, takže DNA konformaci často mírně lišit od B-konformace parametry na Obrázku \(\PageIndex{2}\).,
V prokaryot, DNA se vyskytuje v cytoplazmě (spíše zřejmé, protože neexistuje žádná jiná možnost v těchto jednoduchých organismů), zatímco u eukaryot DNA je uvnitř jádra. Navzdory rozdílům v jejich umístění je úroveň ochrany před vnějšími silami a především jejich velikostí, jak prokaryotická, tak eukaryotická DNA balena s bílkovinami, které pomáhají organizovat a stabilizovat celkovou strukturu chromozomů., Poměrně málo je zřejmé, s ohledem na prokaryotické chromozomální balení ačkoli tam jsou strukturální podobnosti mezi některé z proteinů, nalezené v prokaryotické a eukaryotické chromozomy. Většina úvodních kurzů buněčné biologie se proto drží eukaryotického chromozomálního balení.

nahá DNA, ať už prokaryotická nebo eukaryotická, je extrémně tenký pramen materiálu o průměru zhruba 11 nm. Nicméně, vzhledem k velikosti eukaryotických genomů, pokud by byla DNA uložena tímto způsobem uvnitř jádra, stala by se neovladatelně zamotaná. Obrázek kbelík, do kterého si hodil sto metrů příze, aniž by jakýkoli pokus vůbec organizovat pomocí navíjení nebo seskupování., Nyní zvažte, zda byste být schopni dostat se do kbelíku vytáhnout na jeden pramen, a očekávají, že vytáhnout jen jeden pramen, nebo pokud místo toho budete pravděpodobně muset vytáhnout alespoň malý spleti nití. Buňka dělá v podstatě to, co byste udělali s přízí, abyste ji udrželi organizovanou: je úhledně zabalena do menších, zvládnutelných přaden. V případě, že DNA každého chromozomu je ovinutý kolem histonů komplex tvoří první pořadí chromozomální organizace: nucleosome.
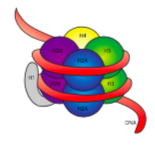
vlákno 30 nm je drženo pohromadě dvěma sadami interakcí. Za prvé, Histon linker, H1, spojuje nukleosomy do přibližné struktury 30 nm., Tato struktura je pak stabilizován disulfidové vazby, které se tvoří mezi histonu H2A jednoho nucleosome a H4 histonů svého souseda.
histony jsou Rodina základních (pozitivně nabitých) proteinů. Všechny funkce především v uspořádání DNA, a nucleosome je tvořen, když DNA zábaly (o něco více než 2 krát) kolem jádra z osmi histony – dva, každý H2A, H2B, H3 a H4. Počet a poloha pozitivních nábojů (většinou z lysinů a argininů) jsou rozhodující pro jejich schopnost pevně vázat DNA, která, jak bylo dříve zdůrazněno, je velmi negativně nabitá., Že“ protiklady přitahují “ myšlenka není jen chodit s někým tip ze sloupců poradenství.

Při vyšetření 3D struktura histonů jádro komplexu, vidíme, že zatímco relativně neutrální protein interakce domén držet histonů spolu v centru města, pozitivně nabité zbytky se nacházejí kolem ven z komplexu, k dispozici na interakci s negativně nabitými fosfáty DNA.,
V pozdější kapitole, budeme diskutovat o tom, jak enzymy přečíst DNA přepsat jeho údaje na menší, lépe zvládnutelné kousky RNA. Prozatím si musíme jen uvědomit, že v daném okamžiku je velká část DNA pevně zabalena, zatímco některé části DNA nejsou. Protože části, které jsou k dispozici pro použití, se mohou lišit v závislosti na tom, co se děje s/v buňce v daném okamžiku, musí být balení DNA dynamické. Musí existovat mechanismus pro rychlé uvolnění vazby DNA na histony, pokud je tato DNA potřebná pro genovou expresi, a pro utažení vazby, Pokud tomu tak není., Jak se ukázalo, tento proces zahrnuje acetylaci a deacetylaci histonů.

Histonů Acetyltransferases (Klobouky) jsou enzymy, které místo acetyl skupiny na lysin a histon bílkovin. Acetylové skupiny jsou negativně nabité a acetylace nejen přidává negativně nabitou skupinu, ale také odstraňuje pozitivní náboj z lysinu. To má za následek nejen neutralizační bod přitažlivosti mezi proteiny a DNA, ale i mírně odpuzující (s poplatky)., Na druhé straně mechanismus, Histonů Deactylases (HDACs) jsou enzymy, které odstraňují se acetylací, a tím obnovení interakce mezi histon bílkovin a DNA. Protože to jsou takové důležité enzymy, je logické, že jim není dovoleno provozovat chtě nechtě na všech dostupných histonů, a ve skutečnosti, oni jsou často nalezené v komplexu s jinými proteiny, které řídí a koordinují jejich aktivace s jinými procesy, jako jsou aktivace transkripce.