
Úvod
Telomery jsou nukleoprotein struktur na velmi koncích lineárních eukaryotických chromozomů. Řeší dva hlavní problémy na konci současně. První je o ochraně konce chromozomů. Odhaduje se, že normální lidské buňky musí opravit alespoň 50 endogenní dvouvláknové zlomy (DSBs) na buňku na buňku-cyklus (Vilenchik a Knudson, 2003)., Telomery odlišují přirozené chromozomální konce od škodlivých DSB a zabraňují jejich ektopické opravě, například konečnými fúzemi chromozomů (vanSteensel a deLange, 1997). Druhým je problém s koncovou replikací, který se zabývá udržováním správných délek telomer. To bylo uznáno nezávisle dvěma vědci (Watson, 1972; Olovnikov, 1973). Od replikativní DNA-dependentní DNA polymerázy nelze dokončit syntézu DNA na velmi koncích chromozomů, kompenzace pro replikační sekvence telomer hmotnosti musí pocházet z RNA-dependentní DNA polymerázy., Tento enzym, zvaný telomeráza, spolu s první telomer minisatellite sekvence, byla objevena v nálevník Tetrahymena (Blackburn a Žlučníku, 1978; Greider a Blackburn, 1985). Jedná se však pouze o jeden aspekt údržby délky telomer. Epigenetické regulaci délky telomer homeostázy, včetně interakce dlouho noncoding telomerické opakovat obsahující RNA a exonuclease aktivity drah, byly také rozsáhle studovány, vzhledem k jeho léčebný potenciál (Wellinger et al., 1996; Polotnianka et al., 1998; Pfeiffer and Lingner, 2012).,
telomeráza, enzym zodpovědný za přidávání sekvencí opakování telomer na 3 ‚ konec telomer, je konzervovaný komplexní enzym s mnoha složkami . V zásadě jen dvě hlavní složky jsou nezbytné pro enzymatickou aktivitu telomerázy, katalyticky aktivní proteinové složky, tzv. telomer reverzní transkriptázy (TERT), a šablony, složka, tvořená telomerasová RNA podjednotku (TR). Zatímco terc je evolučně docela dobře konzervovaný, TR je velmi variabilní, s délkami od ca. 150 nt (Tetrahymena) na více než 2000 nt (houby z rodu Neurospora)., Pouze krátká oblast v celé molekule TR slouží jako šablona pro nově syntetizovanou telomerovou DNA (Greider a Blackburn, 1985; Qi et al., 2013). Tato oblast v TR je obvykle tvořena úplným telomerovým motivem následovaným částečným, který slouží jako žíhání pro stávající telomerovou DNA. Ačkoli je v zásadě zapotřebí pouze jeden další nukleotid (jako částečný motiv), obvykle se nachází více než jeden. Například dva další nukleotidy tvoří žíhací motiv u myší nebo pět u lidí (Blasco et al., 1995; Feng et al., 1995)., U rostlin je však velikost oblasti šablony proměnlivá, např. dvě v Arabidopsis thaliana, sedm v Arabis sp. nebo šest v Nicotianě (Fajkus et al., 2019). Ostatní oblasti TR mají strukturální, regulační a proteinové interaktivní funkce . Viz také schematické zobrazení telomerázy a jejího cyklu aktivity na obrázku 1.

Obrázek 1 Schematické znázornění aktivity telomerázy cyklu s Arabidopsis-typ telomer šablony. Terc, telomer reverzní transkriptáza; TR, podjednotka RNA telomerázy., Obrázek založený na Sekhri (2014).
jak variabilní jsou Telomerové sekvence?
Telomer sekvence jsou obvykle krátké minisatellites tandemly uspořádány, obvykle následující vzorec (TxAyGz)n. Na minisatellite uspořádání pochází od způsobu, ve kterém telomerázy syntetizuje DNA, v krátkosti, a většinou stejné motivy, jeden po druhém., Několik hypotéz domnívají, že takové uspořádání je důležité, protože to podporuje uznání telomer specifických proteinů pomocí homo – a heterodimers a pro potenciál tvaru G-quadruplexes, které mohou stabilizovat koncích chromozomů nebo sloužit jako substráty pro telomer-specifické proteiny (Spiegel et al., 2020; Tran et al., 2013). Telomer sekvence jsou dobře udržovány prostřednictvím evoluce, a velké skupiny organismů pomocí skupiny-typické telomer motivem k vybudování jejich telomer DNA., Postupně rostoucí počet studií a velkých projekcí ukázaly, že všechny testované obratlovce a mnoho bazálních metazoans používat TTAGGG (Meyne et al., 1989; Traut et al., 2007) zatímco Euarthropoda (členovci), včetně Hexapoda (hmyz), mají TTAGG (Frydrychova et al., 2004; Vítková a kol., 2005). Postupně se hromadí četné výjimky, například (A (G)1-8) v Dictyostelium (Emery a Weiner, 1981), TTAGGC v Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG in Coleoptera (brouci) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGGG v Giardii (diplomonads) (Uzlikova et al.,, 2017) nebo TTNNNNAGGG v Yarrowia clade (kvasinky) (Cervenak et al., 2019). Navíc, telomeráza-nezávislé systémy, v nichž minisatellite telomer sekvence byla ztracena a nahrazena komplexní opakuje, jsou zastoupeny, například tím, Dvoukřídlých a Pakomára (přezkoumána v (Mason et al., 2016)). Pro obecný přehled o eukaryotické telomere sekvence viz (Fajkus et al ., 2005; Fulnecká et al., 2013).
složení telomer v rostlinách je ještě rozmanitější., Zde používáme termín „zařízení“ v širším smyslu, také známý jako Archaeplastida či království Plantae sensu lato, a zahrnující Rhodophyta (červené řasy), Glaucophyta, Chlorophyte řas třídy a Streptophyte řas třídy (celkem známé jako zelené řasy), a Embryophyta (půdy, rostliny) (Jeden Tisíc Rostlin Transcriptomes Iniciativy, 2019). Typické rostlinných telomer sekvence je TTTAGGG, také volal Arabidopsis-typ (nebo jednoduše, rostlina-typ), protože byla objevena v Arabidopsis thaliana (Richards a Ausubel, 1988) a nyní v mnoha jiných druhů napříč téměř všechny rostliny objednávky., Ačkoli tttaggg je stále nejčastější, existuje významná variabilita telomerových sekvencí v červených a zelených řasových liniích. Jako pro červené řasy (Rhodophyta), telomer sekvence informace je většinou chybějící nebo neúplné, i když některé telomer kandidátů byly objeveny in silico, jako AATGGGGGG pro Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T) AGGG pro Galdieria sulfuraria (Fulneckova et al., 2013); TTAGGG byl nalezen v genomickém čtení Porphyra umbilicalis (Fulneckova et al., 2013), ale je zapotřebí více důkazů k potvrzení jejich konečné polohy na chromozomech., Rozmanitost telomer v zelených řasách odráží jak dynamické změny, tak jeho paraphyletický charakter. Ačkoli TTTAGGG převládá v Chlorofytech, například v rodech Ostreococcus (Derelle et al., 2006) a Chlorella (Higashiyama et al., 1995), bylo zde také zjištěno mnoho dalších divergentních motivů, jako je TTAGGG v rodu Dunaliella a Stephanosphaeria (Fulneckova et al., 2012) a TTTTAGGG v Chlamydomonas (Petracek et al., 1990). V bazální Streptofytě (Klebsormidiophyceae) byly popsány progresivní změny motivů z TTTAGG na TTTAGG a TTTAGG., Přítomnost TTAGGG v Rhodophyta a Glaucophyta vede k hypotéze, že to je rodový motiv rostliny (Archaeplastida) (Fulneckova et al., 2013).
O suchozemské rostliny, jeden z prvních projekcí provádí ukázal, že Arabidopsis-typ sekvence byl nejčastějším a většinou zachovány prostřednictvím jejich fylogeneze (Cox et al., 1993; Fuchs et al., 1995), ačkoli někteří z těchto autorů již zjistili několik výjimek v rodině Amaryllidaceae (bývalé Alliaceae), ve které sekvence typu Arabidopsis chyběla u několika druhů., Později, první telomer sekvence neobvyklé, půdy, rostlin, obratlovců-typ TTAGGG, byl charakterizován v Aloe vera a v některých dalších Asparagales (Weiss a Scherthan, 2002; Puizina et al., 2003; Sýkorova et al., 2003c). Byla formulována hypotéza o opakovaných ztrátách a výtěžcích sekvence tttaggg a TTAGGG telomere v Asparagales (Adams et al., 2001). S postrefinem řádu Asparagales v APGIII (Angiosperm fylogeneze Group 2009) (Bremer et al.,, 2009), bylo prokázáno, že pouze dvě hlavní evoluční přepínače v telomer sekvence složení došlo (spíše než několik opakované ztráty a zisky), a to v následujícím pořadí: první v rodině, Iridaceae, ve kterém posun ze závodu-typ TTTAGGG na obratlovců-typ TTAGGG se stalo, následuje rodiny Xeronemataceae, Asphodelaceae a jádro Asparagales (včetně Amarillidaceae s.l a Asparagaceae s.l.,); a druhý v rámci podčeledi Allioideae (dříve zacházeno jako samostatné rodiny, Alliaceae), v němž zcela nový telomer sekvence se objevily, CTCGGTTATGGG (Fajkus et al., 2016). Mimo Asparagales, nové telomer sekvence byly také detekovány v půdě rostlina různorodé skupiny jako (i) Solanaceae, ve kterém telomer sekvence Cestrum elegans TTTTTTAGGG byl popsán (Sykorova et al., 2003a; Sýkorova et al., 2003b; Peska et al., 2008; Peska et al.,, 2015) a (ii) Lentibulariaceae, kde rod Genlisea ukázal pozoruhodnou rozmanitost u některých druhů se vyznačuje Arabidopsis-typ telomer opakuje, zatímco jiní vystavoval prolínají sekvence variant TTCAGG a TTTCAGG (Tran et al., 2015).
i Přes všechny telomer motiv výjimky zjištěny skutečné rozmanitosti v telomerické sekvence v suchozemské rostliny je pravděpodobně značně podhodnoceny. Nedávná publikace (Vitales et al.,, 2017), v nichž screening půdy, rostlinných telomer sekvence byla provedena, zjistil, že telomer sekvence byly známé jen jasně pro méně než 10% druhů a 40% z rodů obsažených v Rostlině rDNA databáze (www.plantrdnadatabase.com), zdroj poskytující molekulární cytogenetiky informace o suchozemské rostliny (Garcia et al., 2012). Souhrn distribuce sekvencí telomer v rostlinách po APG IV (skupina Angiosperm fylogeneze, 2016) (Byng et al., 2016), stejně jako nejnovější rostlinná fylogeneze (tisíc rostlin Transkriptomes Initiative, 2019) se nachází na obrázku 2.,
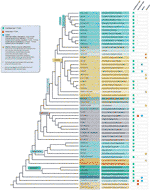
Obrázek 2 Telomer motivy v Archaeplastida (rostliny v širokém slova smyslu), na základě APG IV (Angiosperm Phylogeny Group 2016) a na Jeden Tisíc Rostlin Transcriptomes Iniciativy (2019). Délky větví nevyjadřují váhy v reálném čase., Pro jednoduchost a aby se ušetřilo místo, některé polyphyletic „skupin“ (stupně) označeny hvězdičkou ve stromu byly zastoupeny jedinou pobočku; z téhož důvodu několik menších zakázek (uvedených v modrý čtverec na levé horní straně obrázku) nejsou zobrazeny na strom. První štítek tip obvykle odkazuje na objednávky rostlin a v několika případech na divize, stupně a dokonce i rodiny; druhý štítek zobrazuje reprezentativní rodiny a v několika případech reprezentativní objednávky nebo rody.
od projekcí k objevu: jak lze identifikovat Telomerní motivy?,
důkazy, že daný kandidát pořadí je skutečný telomerické zahrnuje několik kroků, které řádně deklarovat jeho lokalizaci na všech chromozomálních termini, a nakonec zapojení telomerázy v jeho syntézy. Molekulární cytogenetika (většinou pomocí Fluorescenční in situ Hybridizace, FISH) se stala důležitou pro vizualizaci terminálu lokalizace značené sondy kandidáta sekvence na všechny chromozomální termini. Nicméně, samostatné ryby nestačí prokázat samotnou terminální pozici., Například acepsat356 byl navržen v onion jako kandidát telomere na základě výsledků analýzy ryb (Pich a Schubert, 1998). Jeho zdánlivá Poloha terminálu in situ však nikdy nebyla přesvědčivě spojena s funkcí telomere. Ve skutečnosti, objev Allium minisatellite telomer sekvence CTCGGTTATGGG a telomerázy by znamenalo, že AcepSAT356 je subterminal (Fajkus et al., 2019)., Pozitivní RYB telomerické signály mohou také maskovat drobné změny v telomer motivy jako jednonukleotidové polymorfismy, nebo falešně negativní výsledky mohou být důsledkem krátké telomery být pod detekční limit techniky.
existují dva další přístupy, které určují polohu terminálu při větším rozlišení než Ryby; ty jsou založeny na aktivitě exonukleázy BAL31., První je klasické Koncových restrikčních fragmentů (TRF) analýzu, ve které se vzorky zacházeno podle BAL31 show postupné zkracování terminální fragmenty a pokles intenzity signálu s rostoucí čas exonuclease léčby. Následnou analýzu délky fragmentů provádí Southern-blot hybridization (Fojtova et al., 2015). Druhým je komparativní genomové skimming (NGS data) nedigestované a BAL31-trávené genomové DNA, paralelně., V BAL31 léčených dataset, je významným zastoupením sekvence telomer, tedy koncových sekvencí jsou označeny srovnání s neléčenou dataset, pomocí bioinformatických nástrojů RepeatExplorer nebo Tandemových Repetic Finder .
dalším důležitým testem daného kandidáta na sekvenci telomer u druhu je demonstrace aktivity telomerázy. V tomto je užitečným experimentálním přístupem, vyvinutým nejprve pro lidské buňky, protokol Telomere Repeat Amplification Protocol (TRAP) (Kim et al., 1994), následované sekvenováním zjištěných produktů (Peska et al.,, 2015; Fajkus et al., 2016), což je o něco méně citlivé na falešně pozitivní výsledky než ryby. Všechny tyto metody, včetně ryb (Fuchs et al., 1995; Shibata a Hizume, 2011) a další, jako je hybridizace slot-blot (Sykorova et al., 2003c) a TRAP (Fulnečkova et al., 2012; Fulnecká et al., 2016), lze použít k screeningu telomer napříč širokými skupinami složitých organismů, včetně rostlin., Nicméně, pouze kombinace vhodně zvolené metody může přesvědčivě vést k závěru o telomer funkce kandidát pořadí, protože výsledky základny na jeden přístup by mohl být zavádějící. Více kompletní přehled strategií pro de novo telomer kandidát pořadí identifikace, včetně úplně první pokus v Tetrahymena (Greider a Blackburn, 1985) jsou shrnuty v metodické článku, s důrazem na NGS přístupu, který se používá v zařízeních s extrémně velký genom (Peska et al., 2017).
existuje Homoplasie v Telomerových sekvencích?,
rodové sekvence telomer je myšlenka být TTAGGG a je nejvíce běžně vyskytují napříč celým stromem života (Fulneckova et al., 2013). Přesto se zdá být jasné, že frekvence homoplasie v evoluci motivů telomer je relativně vysoká. Například, krátké, jednoduché motivy, jako jsou rostliny-typ TTTAGGG se objevily nezávisle na sobě a opakovaně v cryptomonads, oomycete hub, a alveolates; podobně obratlovců-typ TTAGGG se objevila sekundárně u některých skupin rostlin (Asparagales, Rodophyta a řasy Chlorophyta) (Sykorova et al., 2003c; Fulnecká et al.,, 2012; Fulnecká et al., 2013; Somanathan a Baysdorfer, 2018). Důvod, proč některé telomer sekvence se objevily častěji, než jiné, obvykle složitějších sekvencí je pravděpodobně souvisí s výběrem tlaky, které by si přály přesnost pro konkrétní sekvence specifické DNA-proteinové interakce (Förstermann et al., 2003). Pokud by došlo ke změně v každém telomerovém motivu, zásah do telomerní nukleoproteinové struktury by nutně vedl k nestabilitě genomu., To je důvod, proč telomer sekvence jsou tak evolučně stabilní, obsahující velmi málo nových a úspěšných sekvencí vzoru v souladu s myšlenkou, opakované ztráty a vznik typické telomer sekvence, jak je navrženo pro Asparagales (Adams et al., 2001).
zjištění homoplasie napříč telomerovými sekvencemi vyvolává otázku, jaké jsou molekulární příčiny a procesy probíhající během těchto posunů? Změna telomerní sekvence, přestože se v některých případech jeví jako triviální (např.,, jeden extra T), může způsobit vážné interference s integritou genomu, kvůli narušené rovnováze v interakcích telomer DNA-protein. Není také jasné, zda změna sekvence telomer může mít nějakou evoluční výhodu; v tomto ohledu, (Tran et al., 2015) navrhl, že vzhled „methylatable“ cytosin v G-bohaté telomer pramen by se zvýšila možnost nařízení o epigenetické modifikace.
jaké jsou molekulární důvody změn v motivech telomer?,
pro vysvětlení změny sekvence telomer je prvním kandidátem podjednotka šablony telomerázy, telomerázy RNA (TR). Dříve zjištěné TR z droždí a obratlovců patří do jiné skupiny přepisů, jejichž spojovacím prvkem bylo, že oni byli přepisována pomocí RNA polymerázy II (Pol II)—ale ve všech nálevníků; to bývala jedinou výjimkou z Pol II přepisy před zveřejněním půdy rostlina TR identifikace . Použitím relativně dlouhého telomerového motivu Allium hledat jeho TR v datovém fondu celkové sekvence RNA, Fajkus et al., (2019) ukázal, že dříve vyznačuje noncoding RNA se podílejí na stresové reakci v A. thaliana, tzv. AtR8, byl opravdu telomerasová RNA podjednotku (Wu et al., 2012; Fajkus et al., 2019). Jednalo se o přepis RNA polymerázy III (Pol III) obsahující odpovídající regulační prvky ve své promotorové struktuře. Po dlouhou dobu vědci očekávali, že rostlina TR bude tak odlišná, že by nebylo možné ji identifikovat na základě homologického vyhledávání (Cifuentes-Rojas et al., 2011)., Nicméně, určitý stupeň podobnosti byl úspěšně použit k identifikaci společného TR u několika druhů Allium se srovnávacím výbuchem. Překvapivě, sekvenční homologie, přítomnost stejné regulační prvky, a odpovídající šablony regionu vedla k identifikaci TRs v Allium, Arabidopsis a více než 70 dalších vzdáleně příbuzné rostliny, včetně těch, které se rozcházely telomer motivy jako Genlisea, Cestrum, a Tulbaghia., Pokud víme, stále neexistují žádné údaje o žádném ŘASOVÉM TR, což by objasnilo, zda transkripce TR Pol III je obecnou vlastností všech rostlin nebo ne. Tato práce (Fajkus et al., 2019), založený na CRISPR knock-out a dalších experimentech, také ukázal, že dříve identifikovaný kandidát telomerázové RNA v a.thaliana (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) nebyla funkční šablonu podjednotky telomerázy, jak bylo také prokázáno, krátce po (Dew-Budd et al., 2019)., Za předpokladu, že závislost pol II/Pol III pro transkripci TR je spolehlivým evolučním markerem, budoucí výzkum TR v dalších hlavních eukaryotických liniích pravděpodobně otevře nové poznatky o původu eukaryot. Telomeráza geny a telomer sekvence jsou neuznaných zdrojů informací v tomto směru, a zjištění Pol III závislé TR biogeneze dráhy v nálevník a rostlinné druhy mohou představovat první kroky v tomto směru (Greider a Blackburn, 1989; Fajkus et al., 2019).
jak se chromozomy staly lineárními?,
drtivá většina prokaryot obsahuje kruhové chromozomy, zatímco lineární chromozomy jsou pravidlem v eukaryotách., Proto existují dva možné scénáře, ve kterých buď (i) linearizace byla provedena primitivní telomerázy, předchozí další procesy, které vedly k současné lineární chromozomální vlastnosti a funkce, nebo (ii) linearizace pre-eukaryotické kruhový chromozom byl původně telomerázy nezávislé, ale těsně předtím, než současných eukaryot rozcházely, primitivní telomerázy začali okupovat koncích chromozomů a stala zásadní pro nově vytvořené lineární chromozomy (Nosek et al., 2006). Villasante et al., (2007) navrhuje evoluční scénář, ve kterém dojde k rozbití rodových prokaryotické kruhový chromozom aktivní provedení mechanismu na konce DNA, což umožňuje vznik telomer pomocí rekombinace-dependentní mechanismus replikace: důsledky této hypotézy vedlo k překvapivému závěru, že eukaryotické centromery byly odvozeny od telomery.
Zajímavé je, že opačný proces linearizace, tj. vytvoření kruhové chromozomy (také nazývané ring chromozomy) se objevil čas od času v průběhu evoluce eukaryot, i když je vysoce nestabilní., Například, v případě Amaranthus tuberculatus, ring chromozomy objevil jako stres-indukované reakce, nesoucí rezistenci proti herbicidům (glyfosát); tyto další ring chromozomy neukázal hybridizace s telomer sondy karyotyp analýza (Koo et al., 2018). Téměř univerzální telomerázový systém a výjimečnost kruhových chromozomů v eukaryotách nám neumožňují podporovat jednu hypotézu nad druhou., Rekombinační stroje používané při alternativním prodlužování telomer (ALT), cesty nezávislé na telomeráze, spojené s některými lidskými rakovinami (Zhang et al., 2019), je již přítomen v prokaryotách. Kromě toho existují důkazy o chromozomových linearizace vyskytující se nezávisle na sobě v různých prokaryote linií (Ferdows a Barbour, 1989; Nosek et al., 1995; Volff a Altenbuchner, 2000). Proto se zdá pravděpodobnější hypotéza, že první lineární eukaryotický chromozom (pocházející z předka prokaryota) byl nezávislý na telomeráze., Tam jsou některé příklady, které ukazují, že telomeráza-based systém není nezbytný pro udržování telomer ve všech eukaryot: retrotransposons v Drosophila telomery, satelitní opakuje v Chironomus, jiný hmyz (Rubin, 1978; Biessmann a Mason, 2003), a ALT v telomerázy-negativní lidských nádorů (Hu et al., 2016; Zhang et al., 2019). Přesto, některé z těchto systémů nemusí být tak různé, a možná mají společný původ: v Drosophila, telomer údržbu, se sídlem v retrotransposition, není příliš odlišné od telomerázy-založený mechanismus (Danilevskaya et al.,, 1998), což vede k hypotéze, že samotná telomeráza může být bývalým retrotransposonem. Ale jistě, telomeráza-negativní druhy rostlin nebyly objeveny datum a všechny výjimky, v nichž typické rostliny-typ telomer byl přítomen, bylo později prokázáno, že mají jiný, ale přesto telomerázy-syntetizovány, motivy. Alt strojní zařízení je však přítomno v závodech paralelně s aktivitou telomerázy (Watson a Shippen, 2007; Ruckova et al., 2008). Z toho vyplývají zajímavé otázky týkající se role telomerázy, telomer a jejich udržování v rostlinných nádorech., Atraktivní je o nepřítomnosti metastáz v rostlinách, a to navzdory přítomnosti ALT, snad v souvislosti s rostlinné tkáně tuhost nebo jiný imunitní systém než u zvířat (Seyfried a Huysentruyt, 2013).
přestože získáváme stále větší znalosti o biologii telomer, stále nejsme schopni vysvětlit vznik telomerázy v eukaryotách., Aktuální důkazů podporuje hypotézu, že vznik eukaryot spolu s jejich lineární chromosomy, telomery a telomeráza byla týkající se vzhledu spliceosomal intronů v archaeal hostí (Koonin, 2006; Fajkus et al., 2019). Podobnost mezi TERCEM a jinými retroelementy byla diskutována již nějakou dobu (Pardue et al., 1997). Pozoruhodné je, že poměrně nedávná studie ukázala, že TERC, jako pravděpodobný člen progeny group II intronů, je postupně blízkosti Penelope-jako prvek retrotransposons (Gladyshev a Arkhipova, 2007)., Terc je však pouze jednou ze dvou základních složek telomerázy a TR je ve svém původu ještě záhadnější kvůli nízké sekvenční ochraně ve všech eukaryotách .
závěr
na začátku éry rostlinné genomiky byla sekvence telomer považována za téměř neměnnou. Obecné ochraně telomer a telomerázy systém navrhl, že všechny rostliny mohou mít TTTAGGG rostlin-typ telomer. Identifikace neobvyklých telomerových sekvencí v komplexních rostlinných genomech, v mnoha případech s obrovskými hodnotami C (například v Cestrum a Allium sp.,), stálo za to úsilí, protože mimořádně dlouhý motiv Allium telomere byl vodítkem při hledání skutečného TR v pozemních rostlinách. Nově popsaný TR v rostlinách a dále telomer/telomerázy výzkum v bazálních skupin řas může odhalit cenné informace o raném vývoji, proto rostlinných telomer výzkum může významně přispět k hypotézy na vznik eukaryot.
autorské příspěvky
VP a SG přispěly stejně k psaní, editaci a přípravě této mini-recenze.,
financování
střet zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
iniciativa tisíc Transkriptomů rostlin (2019). Tisíc rostlinných transkriptomů a fylogenomika zelených rostlin. Nature 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstract / CrossRef Full Text / Google Scholar