
Úvod
Nozokomiálních infekcí způsobených multirezistentní Acinetobacter baumannii jsou stále častější na celém světě, zejména v intenzivní péči (Wieland et al., 2018). Úspěch této bakterie je usnadněno jeho schopnost přežít v různých prostředích umocněn jeho rychlá schopnost získat multirezistence. Povrchové sacharidy hrají klíčovou roli v celkové kondici a virulenci a., baumannii (Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Weber et al., 2016). A. baumannii produkuje kapsulární polysacharid s vysokou molekulovou hmotností (CPS), který obklopuje vnější membránu (Obrázek 1) (Russo et al., 2010). Skládá se z pevně zabalené opakování oligosacharid podjednotek (K jednotkám), CPS tvoří diskrétní vrstvu na bakteriální povrch poskytuje ochranu před různorodých podmínek prostředí, pomoc při vyhýbání hostitele imunitní obranu, a zvyšuje odolnost k řadě antimikrobiálních sloučenin (Russo et al., 2010; Iwashkiw et al., 2012; Geisinger and Isberg, 2015).,
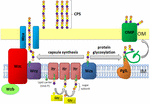
Obrázek 1. Schematické znázornění polysacharidu kapsle sestavení a vývozu v a. baumannii. Syntéza začíná s počáteční transferázy (Itr; maroon) se nachází ve vnitřní membráně (IM) přenos prvního cukr opakující se jednotky na lipidový nosič (Und-P; zelený kruh). Následné cukry se pak přidávají do rostoucí jednotky specifickými glykosyltransferázami (GTR; žlutá) na cytosolické straně vnitřní membrány., Podjednotka kapsle (jednotka K) se pak přenese do periplazmy přes integrální membránový protein Wzx (Tmavě modrá). Cukr podjednotky jsou polymeruje na Wzy bílkovin (fialová) a Wza/Wzb/Wzc komplex (azurová, limetková, červená) souřadnice vysoké úrovni polymerace a vývoz rostoucí řetězce, jejich přesunutí na vnější membrána (OM). Pro glykosylaci pgll (oranžová) spojuje jednotky K na vybrané proteiny vnější membrány (OMP; tmavě zelená).
V a.baumannii dochází k montáži a exportu kapslí cestou závislou na Wzy (Hu et al.,, 2013; Kenyon and Hall, 2013; Willis and Whitfield, 2013; Woodward and Naismith, 2016) (Obrázek 1). Obvykle se skládá z 4-6 cukrů, K jednotce je sestaven na lipidový nosič molekuly undecaprenyl pyrofosforečnan (A-P), který poskytuje lešení pro rostoucí cukr řetězce (Whitfield, 2006). První cukr v K jednotce, je rekrutován vnitřní membrána (IM)-vázané počáteční transferázy (Itr), následuje postupné přidávání cukrů do rostoucí K jednotce konkrétní glykosyl transferázy (Gtr) enzymy (Obrázek 1) (Woodward a Naismith, 2016)., Každý K jednotce je pak převedena na periplasmic straně IM kterou Wzx translocase a polymerované za Wzy, která přenáší rostoucí polysacharidové řetězce z jednoho Und-P dopravce na další příchozí podjednotku (Obrázek 1) (Collins et al., 2007). Po CPS polymer je syntetizován, je transportován k povrchu buňky prostřednictvím vysoce koordinovaný proces, zahrnující interakce tří proteinů; Wza, Wzb, a Wzc, které tvoří vývoz strojů (Obrázek 1)., CPS syntéza představuje jednu paži rozdvojená cesta, jak tyto K jednotky jsou také použity k výzdobě některých povrchových proteinů pomocí O-vázaných proteinů glykosylace (Lees-Miller et al., 2013). V tomto případě jsou jednotlivé jednotky K přeneseny na přijímající proteiny o-oligosacharyltransferázou PglL (Obrázek 1) (Iwashkiw et al., 2012). V. A. baumannii, glykosylace bílkovin přispívá k biofilm vznik zvýšením počáteční uchycení a zrání biofilmy, a patogenita prokázána na mnoha zvířecích infekce modely (Iwashkiw et al., 2012; Scott et al., 2014; Harding et al.,, 2015). Biofilm je růstový stav, ve kterém jsou bakteriální komunity uzavřeny v exopolysacharidové matrici a bylo prokázáno, že hrají významnou roli v perzistenci a rezistenci a.baumannii.
Další povrchové sacharidy je známo, že ovlivňují patogenitu a. baumannii patří lipooligosaccharide (LOS) a exopolysaccharide poly-β-(1-6)-N-acetylglukosamin (PNAG) (Preston et al., 1996; Weber et al., 2016). PNAG tvoří soudržné „lepidlo“ biofilmů a tvoří podstatnou část biofilmů (Choi et al., 2009; Longo et al., 2014)., Na rozdíl od většiny Gram-negativní bakterie A. baumannii nevytváří tradiční lipopolysacharid, ale místo toho podobný povrch glyko-konjugát, LOS, která se skládá z lipidové jádro, které postrádá O-antigen (Kenyon a Hall, 2013; Kenyon et al., 2014b). Ztráta produkce LOS V a. baumannii snižuje stabilitu vnější membrány vedoucí ke snížení kondice (Moffatt et al., 2010; Beceiro et al., 2014).
ačkoli mnoho sacharidů odrůdy ovlivňují patogenitu, lze tvrdit, že CPS je převládající virulence faktor a.baumannii. Cílem tohoto přezkumu je konsolidovat to, co je známo o a., baumannii kapsle včetně vybrané struktury, biosyntézy a genové organizace, role CPS ve virulenci, a potenciál pro CPS jako cíl pro budoucí vakcíny a vývoj léčiv.
Genetické Organizace K Loci
Jako kompletní genomy více A. baumannii izolátů k dispozici, pravda rozmanitost kapsle struktur přítomných v této bakterie je stále patrné. K dnešnímu dni bylo v a.baumannii (obrázek 2a) (Shashkov et al identifikováno více než 100 unikátních kapslí loci (KL)., 2017). Tyto oblasti se obvykle pohybují od 20 do 35 kb., Analýza genů, režie CPS syntézy v deset úplných A. baumannii genomů původně vyústil v označení devět kapsle typy, KL1–KL9, která se stala základem pro univerzální psaní schéma pro tyto lokusy (Kenyon a Hall, 2013). Tento systém se následně rozšířil tak, aby vyhovoval identifikaci nových k loci. Chromozomální umístění k lokusu mezi geny fkpA a lldP je vysoce konzervováno mezi kmeny a.baumannii a obsahuje ty geny potřebné pro biosyntézu a export specifické pro každý typ CPS (Obrázek 2) (Hu et al.,, 2013; Kenyon and Hall, 2013). Výjimkou z tohoto pravidla je A. baumannii kmeny mají KL19 a KL39 regionech, kde je gen kódující Wzy polymerázy, wzy, se nachází na malé genetické ostrova jinde na chromozomu (Kenyon et al., 2016a). Kromě toho se geny potřebné pro některé běžné cukry pozorované v CPS nacházejí jinde., Všechny K loci vykazují podobné genetické konfigurace, vysoce variabilní clusteru syntézy a transferázy geny nutné pro biosyntézu unikátní KL-typ složité cukry, lemovaný na jedné straně vysoce konzervovaným CPS export genů, a na druhé straně o soubor genů kódujících zachovány jednoduché cukry a prekurzorů (Obrázek 2A)., Na wzx a wzy genů potřebných pro opakování-jednotky zpracování jsou vysoce variabilní mezi K loci (Obrázek 2A, světle modrá), což naznačuje specifičnost pro zvláštní jednotka K struktur a obecně, aby gtr determinanty, kódování specifické glycosyltransferases v KL regionů, nepřímo odpovídá s cílem akce.
proměnná regionu, některé KL genové shluky, jako KL37 a KL14, nedostatek genů pro komplexní syntézu cukru, protože obsahují pouze jednoduché cukry v jejich K jednotek (Arbatsky et al., 2015; Kenyon et al.,, 2015). Také přidáním k jejich rozmanitosti obsahuje několik oblastí KL redundantní geny, které nejsou nutné pro syntézu konečné jednotky K. Například KL8 a KL9 obsahují dva itr geny (Kenyon a Hall, 2013) a KL37 má pgt1 phosphoglyceroltransferase, ale žádné odpovídající phosphoglycerol reziduí ve stanovené struktuře (Arbatsky et al., 2015). Kromě toho v KL93 dva prvky vkládací sekvence (ISAba26 a ISAba22) přerušují determinant pgt1 (Obrázek 2) (Kasimova et al., 2017)., Geny potřebné pro specifickou biosyntézu cukru nebudou diskutovány, protože tyto cesty byly dříve pokryty a jsou mimo rozsah tohoto přezkumu (Hu et al., 2013; Kenyon and Hall, 2013). Kromě toho se předpokládá, že další geny umístěné v produktech KL kódují acetylaci nebo acylační modifikaci specifických glykanů (obrázek 2A, růžová). Přestože vyšetření k loci může odhalit hodně o jednotkových strukturách K, je nutná chemická analýza a biochemické testování k určení přesných struktur a identifikaci specifických vazeb mezi opakovanými cukry.,
CPS Konstrukcí
prvních studií na CPS sérotypizaci A. baumannii byly motivovány potřebou vyvinout metodu k diskriminaci. A. baumannii izoluje od ostatních druhů Acinetobacter, stejně jako fenotypová analýza byla zatížena nejednoznačnosti a chybné identifikace (Traub, 1989). Jak je popsáno výše, tam je fenomenální rozmanitost vidět v a. baumannii CPS biosyntézy genových klastrů, což se promítá do rozmanitosti vidět v struktuře jednotky K (Hu et al., 2013; Kenyon and Hall, 2013). Kolektivně, přes 40 různorodé a., struktury jednotek baumannii K byly dosud objasněny pomocí NMR spektroskopie a chemické analýzy. K jednotkové struktury se liší složením cukru. Mohou zahrnovat deriváty běžných cukrů vázaných na UDP, jako je glukóza, galaktóza a kyselina glukuronová nebo vzácné a atypické cukry, jako jsou kyseliny jiné než 2-ulosonové. Struktury se liší délkou a mohou se skládat pouze ze dvou zbytků, jak je vidět u K53 typu CPS (Shashkov et al., 2018), nebo až pět nebo šest monosacharidů, jako je tomu u K37 (obrázek 2b) (Arbatsky et al., 2015)., Struktury se také liší v vazbách jak uvnitř, tak mezi jednotkami K, což vede k výrobě jednotek K, které jsou lineární nebo zahrnují boční větve, jak je vidět v K1 a K93 (obrázek 2b) (Kenyon et al., 2016a; Kasimova et al., 2017). Rozdíly v umístění specifických glykosidických vazeb a o-acetylačních vzorců různých oligosacharidů ve struktuře také přispívají k jednotkové rozmanitosti k.,
Rozdíly mezi K jednotce struktury mohou být jemné, například, K12 a K13 liší pouze tím, že spojení dvou glykany, které vyžaduje použití alternativní Wzy polymerázy; v souladu s tím, K loci oba kmeny jsou totožné, s výjimkou wzy genu (Obrázek 2). Případně změna může být zarážející, jako je začlenění vzácné cukry, včetně pseudaminic, legionaminic, nebo acinetaminic kyseliny deriváty, jak je vidět v K2/6, K49, a K12/13 struktury, respektive (Obrázek 2) (Kenyon et al., 2014a, 2015; Vinogradov et al., 2014; Kasimova et al., 2018)., Zajímavé je, že deriváty kyseliny acinetaminové byly identifikovány pouze v a.baumannii a nenacházejí se nikde jinde v přírodě (Kenyon et al., 2017). Navíc, některé K jednotkám začlenit jedinečný deriváty konkrétní glykany, například pseudaminic kyseliny K93 je acetylovaný s (R)-3-hydroxybutanoyl skupiny (Kasimova et al., 2017), zatímco u k2 a K6 není kyselina pseudaminová acetylována (Obrázek 2)., Kromě toho, struktura K4 je jedinečná tím, že obsahuje pouze aminosugars, D-QuipNAc, a jeden terminál N-acetyl-D-galactosamine (D-GalpNAcA) pobočka, která je limitován s pyruvyl skupiny, vzácný motiv a první budou popsány v Acinetobacter (Obrázek 2A, černá) (Kenyon et al., 2016b). Jako počet K jednotce struktury objasněn zvyšuje, takže dělá jistotou odvodit K jednotce struktury z analýzy, biosyntézy klastry umístěné v A. baumannii KL., Ačkoli je informativní, pochopení role CPS hraje v patogenezi, je důležité aplikovat tyto znalosti na zlepšení výsledků infekcí a. baumannii.
Role ve virulenci, antimikrobiální rezistenci a perzistenci
není pochyb o tom, že přítomnost CPS je nezbytná pro patogenitu a. baumannii. Nejen, že je nezbytné pro vyhýbání se hostitelské imunitní obrany (Russo et al., 2010; Geisinger a Isberg, 2015), ale je důležité pro odolnost vůči antimikrobiálním sloučeninám a přežití v nepříznivých prostředích (Luke et al., 2010; Russo et al.,, 2010; Geisinger and Isberg, 2015). CPS zprostředkovává imunitní úniky v mnoha A. baumannii kmeny omezením interakce mezi imunogenní povrchové struktury bakterií a hostitelskou obranu (Preston et al., 1996; Wu et al., 2009; Russo et al., 2010; Umland et al., 2012; Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Wang-Lin et al., 2017). Zrušení kapsle u více různých kmenů a. baumannii ukázalo snížené přežití v lidském séru a ascites tekutině a útlum u modelů infekce potkanů a myší (Russo et al., 2010; Umland et al., 2012; Lees-Miller et al.,, 2013; Sanchez-Larrayoz et al., 2017). Kromě toho, že up-regulace kapsle výroby v běžně využívána A. baumannii kmen ATCC 17978 (K3 CPS typ) zvýšené sérové rezistenci a virulenci v myším modelu infekce (Geisinger a Isberg, 2015). Kromě toho by mohla být vyvinuta nová antimikrobiální léčba pro specifické typy CPS, například ty, které obsahují kyselinu pseudaminovou, protože její přítomnost byla korelována se zvýšenou virulencí (Hitchen et al., 2010; Kao et al., 2016).
kromě ochrany před hostitelskou obranou, v A., baumannii CPS produkce zvyšuje odolnost k řadě antimikrobiálních sloučenin, včetně těch, které se používají pro dezinfekci v klinických nastavení (Geisinger a Isberg, 2015; Tipton et al., 2015; Chen et al., 2017). Kromě toho růst a. baumannii v subinhibičních hladinách antimikrobiálních látek ovlivňuje produkci CPS. Například, působení antibiotika, chloramfenikol nebo erytromycin vedlo k rozšířené tobolka syntézy v ATCC 17978 (Geisinger a Isberg, 2015) a meropenem expozice pro vybrané mutace vedoucí ke ztrátě v CPS výroby v izolátu 37662 (Chen et al., 2017)., Studie provedené na širší škále kmenů a. baumannii jsou nutné k určení, zda je ochrana proti antimikrobiálním látkám poskytovaná CPS specifická pro kmen, specifická pro kapsle nebo univerzální.
schopnost A. baumannii přetrvávat v klinickém prostředí má nepochybně lepší kolonizace a infekce u citlivých pacientů. A. baumannii je schopen přežít měsíce na nemocničních površích, jako jsou kolejnice, nábytek a zdravotnické prostředky, které poskytují rezervoár, který je často zdrojem přenosu a infekce (Wendt et al., 1997; Gayoso et al., 2013)., Pozoruhodné tolerance vysychání a. baumannii je myšlenka být kvůli „poprsí nebo boom“ strategie, kde přetrvávající subpopulace buněk přežít na úkor umírajících buněk; CPS zvyšuje vysušení tolerance tím, že poskytuje fyzickou bariéru usnadňuje zadržování vody (Roberts, 1996; Webster et al., 2000; Gayoso et al., 2013; Bravo et al., 2016). Přímá role CPS v odolnosti proti vysychání byla nedávno prokázána u kmene a. baumannii AB5075 (Typ K25 CPS). V této studii byla akapsulární varianta AB5075 zobrazena 2.,5-násobné snížení životaschopnosti ve srovnání s rodičovským kmenem(Tipton et al ., 2018). Navíc, ve dvou blízkých příbuzných. A. baumannii, Acinetobacter calcoaceticus a Acinetobacter baylyi, výroba exopolysaccharide a/nebo CPS bylo prokázáno, že podporuje vysychání přežití (Roberson a Firestone, 1992; Ophir a Gutnick, 1994). Kromě ovlivnění odolnosti proti vysychání byl CPS spojen s dalšími virulenčními rysy včetně motility (McQueary et al., 2012; Huang et al., 2014) a výroba biofilmu (Umland et al., 2012; Lees-Miller et al.,, 2013), čímž upevňuje svou roli patogenního faktoru.
nedávné studie spojily fázově variabilní fenotyp a.baumannii AB5075 se změnami ve výrobě CPS, protože vysoce virulentní neprůhledné varianty vytvářejí vrstvu CPS s dvojnásobnou tloušťkou jejich průsvitných protějšků (Chin et al., 2018). Tento přechod z průsvitného na neprůhledný také dramaticky zvýšil patogenní potenciál a. baumannii AB5075. Byla také zvýšena odolnost vůči běžným nemocničním dezinfekčním prostředkům a podmnožině aminoglykosidových antibiotik (Tipton et al., 2015; Chin et al.,, 2018) a neprůhledné varianty byly také více odolné vůči lidské lysozym, cathelicidin souvisejících s antimikrobiální peptid LL37 a peroxidu vodíku ve srovnání s průsvitné kolonie (Brady et al., 2018). Kromě toho měly neprůhledné izoláty zvýšenou toleranci vůči vysušeným podmínkám a out-soutěžily průsvitné protějšky v modelu infekce myší (Chin et al., 2018). Jako více faktory jsou zapojeny ve fázi variace, další studie byly provedeny pro stanovení příspěvku CPS výroby do více virulentní neprůhledné fenotyp., V následující publikaci autoři prokázali, že akapsulární varianta byla významně náchylnější k lysozymu a dezinfekčním prostředkům ve srovnání s neprůhledným rodičem divokého typu AB5075 (Tipton et al., 2018). Zajímavé je, že nebyl žádný rozdíl v odolnosti vůči LL-37 a peroxid vodíku mezi neprůhledné wild-type a acapsular kmenů, což naznačuje, že jiné faktory než CPS výroby vedou k tomuto fenotypu pro neprůhledné varianty a. baumannii (Tipton a kol., 2018).,
Nařízení CPS Výroby
Environmentální podněty, jako je teplota, osmotický tlak a změny v metabolitů a iontů dostupnost může ovlivnit bakteriální CPS výroby (Hagiwara et al., 2003; Lai et al., 2003; Mouslim et al., 2004; Willenborg et al., 2011). Je překvapující, že několik regulačních mechanismů byly identifikovány pro CPS výroby jako CPS úrovně jsou běžně regulovány post-translationally prostřednictvím fosforylace CPS export strojů (Whitfield a Paiment, 2003; Chiang et al., 2017). V A., baumannii, pouze dva regulátory kapsle výroby dosud nebyly identifikovány; BfmRS a OmpR-EnvZ, obě dvě složky signálu transdukční systémy, které hrají více regulační úlohu v obálce biogeneze (Geisinger a Isberg, 2015; Tipton a Spíše 2017; Geisinger et al., 2018). Když se podrobí antibiotikum tlak, A. baumannii ATCC 17978 cps exprese byla zvýšena v BfmRS-závislým způsobem (Geisinger a Isberg, 2015). Fázová variace, a tedy potenciálně výroba CPS, je vysoce regulována systémem OmpR-EnvZ V A., baumannii AB5075, jako mutace v OmpR nebo EnvZ za následek výrazné zvýšení neprůhledné až průsvitné spínací frekvence (Tipton a Spíše 2017). Přestože přechod z průsvitného na neprůhledný vede k dvojnásobnému zvýšení tloušťky kapsle, transkriptomické analýzy nezjistily žádné rozdíly v úrovních exprese genů KL mezi oběma fázemi.
CPS jako cíl pro vývoj vakcín a ošetření proti antibiotikům rezistentním na a. baumannii
(konkrétně cabapenem) a., baumannii jsou klasifikovány jako Světová zdravotnická organizace priorita 1 kritický organismus pro vývoj nových antimikrobiálních látek (WHO, 2017). Ačkoli v současné době neexistují žádné antibiotické léčby nebo vakcíny licencované Pro a. baumannii, existuje zvýšený zájem o jejich vývoj a předběžné studie vypadají slibně. Povrchová expozice a prevalence patogenních kmenů a. baumannii činí CPS ideálním cílem jak pro antimikrobiální ošetření, tak pro vakcíny (Russo et al., 2013)., Mezi ně patří vývoj protilátek-terapie založené jako profylaktické vakcíny, pasivní imunizace, a fágové terapie (García-Quintanilla et al., 2013).
Několik studií, které prokázaly účinnost pasivní imunizace u myší pomocí CPS-specifické protilátky, které je chrání proti bakteriální výzva s 13-55% klinických A. baumannii izolátů (Russo et al., 2013; Yang et al., 2017; Lee et al., 2018)., Navíc očkování s konjugovanou očkovací látky obsahující CPS glykany připojen k proteinu dopravce vyvolat lepší imunitní ochranu, než čištěný CPS proti pestrou škálu A. baumannii kmenů (Yang et al., 2017).
Zájem fágové terapie k léčbě bakteriálních infekcí se v posledních letech zvýšila v reakci na současnou krizi rostoucí antimikrobiální rezistence. Terapie fágem je atraktivní jako potenciální léčba infekcí a.baumannii odolných vůči více lékům., Například, phage kódování CPS depolymerase bylo zjištěno, že snížit CPS přibližně 10%, čtyři z 38, klinické multirezistentní A. baumannii testovány (Hernandez-Morales et al., 2018). I když hostitel rozsah tohoto fága je omezené, to může být začleněna jako součást phage koktejl chcete-li maximalizovat efektivitu, nebo cílit na konkrétní ohniska (Hernandez-Morales et al., 2018). Fág, který selektivně štěpí vazbu a., baumannii CPS na pseudaminic kyseliny pobočky mohou být cenné pro fágové terapie, nebo efektivně výrobě čištěné CPS pro vakcíny a protilátky rozvoj (Lee et al., 2018). Phage cílení a. baumannii bylo nedávno prokázáno, že je stabilní, když je impregnován v produktech pro péči o popáleniny za různých podmínek, včetně přítomnosti antimikrobiálních látek (Merabishvili et al., 2017). Nicméně, protože většina studií vyšetřujících a. baumannii CPS je obvykle omezena na určitý kmen a. baumannii, není známo, zda se tato zjištění promítají do všech a., izoláty baumannii nebo pokud jsou specifické pro typ kmene nebo kapsle. Pochopení toho, co role kapsle hraje v několika kmenů je rozhodující při identifikaci CPS typy, které představují to nejlepší cíle pro nové vakcíny, nebo zda vývoj antimikrobiálních látek cílení kapsle biosyntéza cesta je opravdu ještě možné.
Závěrečné Poznámky
i když kapsle představuje důležitý virulence rys A. baumannii jsou dostupné omezené údaje o úloze různých CPS typy hrát v způsobit onemocnění., Vyvinout účinné vakcíny a terapie zaměřené CPS, musíme nejprve získat komplexní pochopení vůči mechanismy za jeho syntézu a výraz, vedle výhod, které kapsle vyjadřuje do hostitelské bakterie. Tento výzkum musí být řešeny v kontextu extrémní variantu CPS sérotypy nalezen v A. baumannii, aby bylo zajištěno, případné zásahy proti kmeny produkující rozmanité CPS struktur., Další studie na CPS jsou povinni poskytnout platformu pro rozvoj preventivní opatření a ošetření proti této stále přetrvávající a smrtící lidský patogen.
autorské příspěvky
JS napsal první návrh. MB poskytl akademický vstup a kritickou revizi článku. FA produkoval zarovnání genomu a za předpokladu, kritickou revizi rukopisu. Všichni autoři schválili konečnou verzi.
financování
tato práce byla podpořena grantem Flinders Medical Research Foundation na MB. FA byl podporován AJ a IM Naylon a Playford Trust Ph.D., Stipendium. JS byl podporován aj a IM Naylon stipendium.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Traub, W. H. (1989). Acinetobacter baumannii sérotyping pro vymezení ohnisek nozokomiální křížové infekce. J. Clin. Mikrobiol. 27, 2713–2716.
PubMed Abstract / Google Scholar
Whitfield, C., and Paiment, a. (2003)., Biosyntéza a sestavení kapsulárních polysacharidů skupiny 1 v Escherichia coli a souvisejících extracelulárních polysacharidech v jiných bakteriích. Karbohydr. Res. 338, 2491-2502. doi: 10.1016 / j. carres.2003.08.010
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
KDO (2017). Globální seznam priorit bakterií rezistentních na antibiotika, které vedou výzkum, objev a vývoj nových antibiotik. Ženeva: Světová Zdravotnická Organizace.
Google Scholar