
como se puede ver en la Figura 1, los nucleótidos solo varían ligeramente, y solo en la base nitrogenada. En el caso del ADN, esas bases son adenina, guanina, citosina y timina. Nótese la similitud de las formas de adenina y guanina, y también la similitud entre citosina y timina. A y G se clasifican como purinas, mientras que C y T se clasifican como pirimidinas. Mientras estemos nombrando cosas, fíjate en «desoxirribosa»y » ribosa». Como su nombre lo indica, la desoxirribosa es solo una ribosa sin oxígeno., Más específicamente, donde hay un grupo hidroxilo unido al 2-carbono de la ribosa, solo hay un hidrógeno unido al 2-carbono de la desoxirribosa. Esa es la única diferencia entre los dos azúcares.
en la construcción aleatoria de una sola hebra de ácido nucleico in vitro, no hay reglas particulares con respecto al ordenamiento de los nucleótidos con respecto a sus bases. Las identidades de sus bases nitrogenadas son irrelevantes porque los nucleótidos están unidos por enlaces fosfodiéster a través del grupo fosfato y la pentosa., Por lo tanto, a menudo se conoce como la columna vertebral de azúcar-fosfato. Si desglosamos la palabra «fosfodiéster», vemos que describe bastante fácilmente la conexión: los azúcares están conectados por dos enlaces éster ( —O—) con un fósforo en el medio. Una de las ideas que a menudo confunde a los estudiantes es la direccionalidad de este enlace, y por lo tanto, de los ácidos nucleicos en general. Por ejemplo, cuando hablamos de la ADN polimerasa, la enzima que cataliza la adición de nucleótidos en las células vivas, decimos que funciona en una dirección de 5-prime (5′) a 3-prime (3′)., Esto puede parecer un lenguaje arcano de biólogos moleculares, pero en realidad es muy simple. Eche otro vistazo a dos de los nucleótidos Unidos por el enlace fosfodiéster (figura \(\PageIndex{1}\), abajo a la izquierda). Un nucleótido de adenina se une a un nucleótido de citosina. El enlace fosfodiéster siempre unirá el 5-carbono de una desoxirribosa (o ribosa en ARN) al 3-carbono del siguiente azúcar. Esto también significa que en un extremo de una cadena de nucleótidos enlazados, habrá un grupo libre de 5′ fosfato (-PO4), y en el otro extremo, un hidroxilo libre de 3′ (- OH)., Estos definen la direccionalidad de una cadena de ADN o ARN.

El ADN se encuentra normalmente como una molécula de doble cadena en la célula, mientras que el ARN es principalmente de una sola cadena., Es importante entender, sin embargo, que bajo las condiciones apropiadas, el ADN podría ser de una sola cadena, y el ARN puede ser de doble cadena. De hecho, las moléculas son tan similares que incluso es posible crear moléculas híbridas de doble cadena con una hebra de ADN y otra de ARN. Curiosamente, las hélices dobles ARN-ARN y las hélices dobles ARN-ADN son en realidad ligeramente más estables que la hélice doble ADN-ADN más convencional.,
la base de la naturaleza de doble cadena del ADN, y de hecho la base de los ácidos nucleicos como medio para el almacenamiento y la transferencia de información genética, es el emparejamiento de bases. Apareamiento de bases se refiere a la formación de enlaces de hidrógeno entre las adeninas y timinas, y entre guaninas y citosinas. Estos pares son significativamente más estables que cualquier asociación formada con las otras bases posibles. Además, cuando estas asociaciones de pares de bases se forman en el contexto de dos hebras de ácidos nucleicos, su espaciamiento también es uniforme y altamente estable., Usted puede recordar que los enlaces de hidrógeno son enlaces relativamente débiles. Sin embargo, en el contexto del ADN, el enlace de hidrógeno es lo que hace que el ADN sea extremadamente estable y, por lo tanto, adecuado como medio de almacenamiento a largo plazo para la información genética. Dado que incluso en procariotas simples, las hélices dobles del ADN tienen al menos miles de nucleótidos de largo, esto significa que hay varios miles de enlaces de hidrógeno que mantienen juntas las dos hebras., Aunque cualquier interacción individual de enlace de nucleótido a nucleótido podría interrumpirse temporalmente por un ligero aumento de la temperatura, o un minúsculo cambio en la fuerza iónica de la solución, una doble hélice completa de ADN requiere temperaturas muy altas (generalmente más de 90oC) para desnaturalizar completamente la doble hélice en hebras individuales.
debido a que hay un emparejamiento uno a uno exacto de nucleótidos, resulta que las dos hebras son esencialmente copias de seguridad una de la otra, una red de seguridad en el caso de que los nucleótidos se pierdan de una hebra., De hecho, incluso si partes de ambas hebras están dañadas, siempre y cuando la otra hebra esté intacta en el área de daño, entonces la información esencial sigue ahí en la secuencia complementaria de la hebra opuesta y puede escribirse en su lugar. Sin embargo, tenga en cuenta que mientras que una hebra de ADN puede actuar como una «copia de seguridad» de la otra, las dos hebras no son idénticas, son complementarias. Una consecuencia interesante de este sistema de hebras complementarias y antiparalelas es que las dos hebras pueden llevar información única.,
los pares de genes bidireccionales son dos genes en hebras opuestas de ADN, pero que comparten un promotor, que se encuentra entre ellos. Dado que el ADN solo se puede hacer en una dirección, 5′ a 3′, este promotor bidireccional, a menudo una isla CpG (véase el capítulo siguiente), envía la ARN polimerasa para cada gen en direcciones físicas opuestas. Esto se ha demostrado para un número de genes implicados en cánceres (pecho, ovario), y es un mecanismo para coordinar la expresión de redes de productos del gen.
las hebras de una doble hélice de ADN son antiparalelas., Esto significa que si miramos una doble hélice de ADN de izquierda a derecha, una hebra se construiría en la dirección 5′ a 3′, mientras que la hebra complementaria se construye en la dirección 3′ a 5′. Esto es importante para la función de las enzimas que crean y reparan el ADN, como hablaremos pronto. En la figura \(\PageIndex{1}\), la cadena izquierda es de 5′ a 3′ de arriba a abajo, y la otra es de 5′ a 3′ de abajo a arriba.
desde un punto de vista físico, las moléculas de ADN están cargadas negativamente (todos esos fosfatos), y normalmente una doble hélice con un giro a la derecha., En este estado normal (también llamado conformación «B»), Un giro completo de la molécula abarca 11 pares de bases, con 0.34 nm entre cada base de nucleótido. Cada una de las bases nitrogenadas son planas, y cuando se empareja con la base complementaria, forma un «peldaño» plano en la «escalera» del ADN. Estos son perpendiculares al eje longitudinal del ADN. La mayor parte del ADN flotante en una célula, y la mayor parte del ADN en cualquier solución acuosa de osmolaridad casi fisiológica y pH, se encuentra en esta conformación B., Sin embargo, se han encontrado otras conformaciones, generalmente bajo circunstancias ambientales muy específicas. Se observó una conformación comprimida, A-DNA, como un artefacto de cristalización in vitro, con un poco más de bases por giro, una longitud de giro más corta y pares de bases que no son perpendiculares al eje longitudinal. Otro, El ADN-Z, parece formarse transitoriamente en tramos de ADN ricos en GC en los que, curiosamente, el ADN se tuerce en la dirección opuesta.
se ha sugerido que las formas A Y Z del ADN son, de hecho, fisiológicamente relevantes., Hay evidencia que sugiere que la forma A puede ocurrir en doble hélices híbridas ARN-ADN, así como cuando el ADN se complexed a algunas enzimas. La conformación Z puede ocurrir en respuesta a la metilación del ADN. Además, la conformación» normal » del ADN-B es algo así como una estructura idealizada basada en estar totalmente hidratada, como es ciertamente muy probable dentro de una célula. Sin embargo, ese estado de hidratación está cambiando constantemente, aunque minuciosamente, por lo que la conformación del ADN a menudo variará ligeramente de los parámetros de conformación B en la figura \(\PageIndex{2}\).,
en procariotas, el ADN se encuentra en el citoplasma (bastante obvio ya que no hay otra opción en esos organismos simples), mientras que en eucariotas, el ADN se encuentra dentro del núcleo. A pesar de las diferencias en sus ubicaciones, el nivel de protección de las fuerzas externas y, sobre todo, sus tamaños, tanto el ADN procariótico como el eucariótico están empaquetados con proteínas que ayudan a organizar y estabilizar la estructura cromosómica general., Se entiende relativamente poco con respecto al empaquetado cromosómico procariótico, aunque hay similitudes estructurales entre algunas de las proteínas que se encuentran en los cromosomas procarióticos y eucarióticos. Por lo tanto, la mayoría de los cursos introductorios de biología celular se adhieren al empaquetado cromosómico eucariótico.

El ADN desnudo, ya sea procariótico o eucariótico, es una hebra extremadamente delgada de material, aproximadamente 11 nm de diámetro. Sin embargo, dado el tamaño de los genomas eucariotas, si el ADN se almacenara de esa manera dentro del núcleo, se enredaría de forma inmanejable. Imagina un cubo en el que has arrojado cien metros de hilo sin ningún intento de organizarlo enrollándolo o agrupándolo., Ahora considere si usted sería capaz de llegar a ese cubo tirar de un hilo, y esperar a tirar de solo un hilo, o si en su lugar es probable que tire hacia arriba al menos una pequeña maraña de hilo. La célula hace esencialmente lo que usted haría con el hilo para mantenerlo organizado: se empaqueta cuidadosamente en madejas más pequeñas y manejables. En el caso del ADN, cada cromosoma se enrolla alrededor de un complejo histónico para formar el primer orden de organización cromosómica: el nucleosoma.
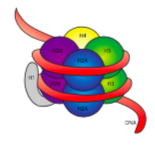
la fibra de 30 nm se mantiene unida por dos conjuntos de interacciones. En primer lugar, la histona enlazadora, H1, reúne los nucleosomas en una estructura aproximada de 30 nm., Esta estructura es estabilizada por enlaces disulfuro que se forman entre la histona H2A de un nucleosoma y la histona H4 de su vecino.
las histonas son una familia de proteínas básicas (cargadas positivamente). Todos funcionan principalmente en la organización del ADN, y el nucleosoma se forma cuando el ADN se envuelve (un poco más de 2 veces) alrededor de un núcleo de ocho histonas, dos de H2A, H2B, H3 y H4. El número y la posición de las cargas positivas (principalmente de lisinas y argininas) son cruciales para su capacidad de unir firmemente el ADN, que como se señaló anteriormente, está muy cargado negativamente., Esa idea de «los opuestos se atraen» no es solo un consejo de Citas de las columnas de consejos.

al examinar la estructura 3D del complejo central de histonas, vemos que mientras que los dominios de interacción de proteínas relativamente sin carga mantienen las histonas juntas en el centro, los residuos cargados positivamente se encuentran alrededor del exterior del complejo, disponibles para interactuar con los fosfatos cargados negativamente del ADN.,
en un capítulo posterior, discutiremos cómo las enzimas leen el ADN para transcribir su información en trozos más pequeños y manejables de ARN. Por ahora, solo necesitamos ser conscientes de que en un momento dado, gran parte del ADN está empaquetado herméticamente, mientras que algunas partes del ADN no lo están. Debido a que las partes que están disponibles para su uso pueden variar dependiendo de lo que está sucediendo a/en la célula en un momento dado, el embalaje del ADN debe ser dinámico. Debe haber un mecanismo para aflojar rápidamente la Unión del ADN a las histonas cuando ese ADN es necesario para la expresión génica, y para apretar la Unión cuando no lo es., Resulta que este proceso implica acetilación y desacetilación de las histonas.

las histonas Acetiltransferasas (HATs) son enzimas que colocan un grupo acetil en una lisina de una proteína histona. Los grupos acetilo están cargados negativamente, y la acetilación no solo agrega un grupo cargado negativamente, sino que también elimina la carga positiva de la lisina. Esto tiene el efecto de no solo neutralizar un punto de atracción entre la proteína y el ADN, sino incluso repelerlo ligeramente (con cargas similares)., En el otro lado del mecanismo, las histonas Deactilasas (HDACs) son enzimas que eliminan la acetilación y, por lo tanto, restauran la interacción entre la proteína histona y el ADN. Dado que estas son enzimas tan importantes, es lógico pensar que no se les permite operar de manera voluntaria en ninguna histona disponible, y de hecho, a menudo se encuentran en un complejo con otras proteínas que controlan y coordinan su activación con otros procesos como la activación de la transcripción.