
Kuten voit nähdä Figure1, nukleotidit vain vaihtelevat hieman, ja vain typpipitoiset base. DNA: n tapauksessa nämä emäkset ovat adeniini, guaniini, sytosiini ja tymiini. Huomaa samankaltaisuus muotoja adeniini ja guaniini, ja myös samankaltaisuuden sytosiini ja tymiini. A ja G luokitellaan puriineiksi, kun taas C ja T luokitellaan pyrimidiineiksi. Kunhan nimeämme asioita, huomaa ”deoxyribose”ja ” riboose”. Kuten nimestä voi päätellä, deoksiriboosi on vain riboosi ilman happea., Tarkemmin, jossa on hydroksyyliryhmä kiinnitetty 2-hiili-riboosi, on vain vetyä kiinnitetty 2-hiilen deoxyribose. Se on ainoa ero näiden kahden sokerin välillä.
satunnaisesti rakentaa yhden lohkon nukleiinihappo in vitro, ei ole olemassa erityisiä sääntöjä, jotka koskevat tilaaminen nukleotidien osalta niiden perusteet. Identiteetit niiden typpipitoiset emäkset ovat merkityksettömiä, koska nukleotidit ovat kiinnitetty phosphodiester joukkovelkakirjojen kautta fosfaatti ryhmä ja pentoo., Siksi sitä kutsutaan usein sokeri-fosfaattirungoksi. Jos me murtaa sana ”phosphodiester”, näemme, että se varsin kätevästi kuvataan yhteys: sokerit on yhdistetty kaksi esteri joukkovelkakirjoja ( —O—), jossa on fosforia välillä. Yksi idea, joka usein hämmentää opiskelijoita on suuntaavuus tämän bond, ja siksi, nukleiinihappoja yleensä. Esimerkiksi, kun puhumme DNA-polymeraasi on entsyymi, joka katalysoi lisäksi nukleotidien eläviä soluja, voimme sanoa, että se toimii 5-prime (5′) 3-prime (3′) suuntaan., Tämä saattaa vaikuttaa arkaaniselta molekyylibiologilta, mutta se on itse asiassa hyvin yksinkertaista. Ottaa toisen tarkastella kaksi nukleotidien liittyi yhdessä phosphodiester sidos (Kuva \(\PageIndex{1}\), alhaalla vasemmalla). Adeniininukleotidi liitetään sytosiinininukleotidiin. Fosfodiesterisidos yhdistää aina yhden deoksiriboosin (tai riboosin RNA: ssa) 5-hiilen seuraavan sokerin 3-hiileen. Tämä tarkoittaa myös sitä, että toinen pää ketjun yhteydessä nukleotidit, siellä on vapaa 5′ – fosfaatti (-PO4) ryhmä, ja toinen pää, vapaa 3′ – hydroksyyli – (- OH)., Nämä määrittelevät DNA-tai RNA-juosteen suuntauksen.

– DNA on yleensä löytynyt kuin kaksinkertaisen-pulaan molekyyli, solu ottaa huomioon, että RNA on enimmäkseen single-pulaan., On kuitenkin tärkeää ymmärtää, että sopivissa olosuhteissa DNA voitaisiin tehdä yksijuosteiseksi, ja RNA voi olla kaksijuosteinen. Molekyylit ovat itse asiassa niin samanlaisia, että on jopa mahdollista luoda kaksijuosteisia hybridimolekyylejä, joissa on yksi DNA-juoste ja yksi RNA. Mielenkiintoista, RNA-RNA-double-heliksejä ja RNA-DNA-double heliksejä ovat itse asiassa hieman vakaampi kuin tavanomainen DNA-DNA-kaksoiskierre.,
perusteella double-stranded luonne DNA, ja itse asiassa perustuu nukleiinihappoja, kuten keskipitkän varastointi ja siirto geneettisen informaation, on base-pariksi. Emäsparituksella tarkoitetaan adeniinien ja tymiinien sekä guaniinien ja sytosiinien välisten vetysidosten muodostumista. Nämä parit ovat huomattavasti vakaampia kuin mikään muiden mahdollisten emästen kanssa muodostettu assosiaatio. Lisäksi, kun nämä base-pari yhdistykset lomakkeen yhteydessä kaksi osa-nukleiinihapot, niiden väli on myös yhtenäinen ja erittäin vakaa., Saatat muistaa, että vetysidokset ovat suhteellisen heikkoja sidoksia. Kuitenkin yhteydessä DNA -, vety liimaus on mitä tekee DNA-erittäin vakaa ja siksi sopii hyvin pitkän aikavälin varastointia keskisuurten geneettisen tiedon. Koska jopa yksinkertainen prokaryootit, DNA double heliksejä ovat vähintään tuhansia nukleotidien pitkä, tämä tarkoittaa, että on olemassa useita tuhatta vetysidokset tilalla kaksi säiettä yhteen., Vaikka yksittäisen nukleotidi-to-nukleotidin vety liimaus vuorovaikutus voisi helposti olla tilapäisesti häiritsi hieman lämpötilan nousu, tai minimaalisia muutoksia ionic vahvuus ratkaisu, täysi double-helix DNA vaatii erittäin korkeita lämpötiloja (yleensä yli 90oC) täysin denaturoida double helix yksittäisiä säikeitä.
Koska siellä on tarkka yksi-yhteen pariksi nukleotidien, se kääntyy pois, että kaksi säikeet ovat pääosin varmuuskopio toistensa kopioita – turvaverkko siinä tapauksessa, että nukleotidit ovat menettäneet yhden lohkon., Itse asiassa, vaikka osat molemmat säikeet ovat vaurioituneet, niin kauan kuin toinen säie on ehjä alueella vahinko, niin olennainen tieto on edelleen olemassa täydentäviä järjestyksessä vastakkaisen lohkon ja voi olla kirjoitettu osaksi paikka. Muista kuitenkin, että vaikka toinen DNA-juoste voi siten toimia toisen ”varana”, nämä kaksi juostetta eivät ole identtisiä – ne täydentävät toisiaan. Mielenkiintoinen seuraus tästä täydentävien ja antiparallel-säikeiden järjestelmästä on se, että nämä kaksi osa-aluetta voivat kukin kuljettaa ainutlaatuista tietoa.,
kaksisuuntaiset geeniparit ovat kaksi geeniä DNA: n vastakkaisilla säikeillä, mutta jakavat promoottorin, joka on niiden välissä. Koska DNA voidaan tehdä vain yhteen suuntaan, 5′ 3′, tämä bi-suuntaava promoottori, usein CpG-saarelle (ks. seuraava luku), mikä lähettää RNA-polymeraasi kunkin geenin vastakkaisiin fyysinen suuntiin. Tämä on osoitettu useille syöpiin (rinta -, munasarja -) osallistuville geeneille, ja se on mekanismi geenituotteiden verkostojen ilmentymisen koordinoimiseksi.
DNA: n kaksoiskierteen säikeet ovat antiparallelia., Tämä tarkoittaa, että jos me katsoimme double-helix DNA vasemmalta oikealle, yksi säie olisi rakennettu 5′ 3′ suuntaan, kun täydentäviä strand on rakennettu 3′ 5′ suuntaan. Tämä on tärkeää DNA: ta synnyttävien ja korjaavien entsyymien toiminnalle, sillä keskustelemme asiasta pian. Kuvassa \(\PageIndex{1}\), vasen strand on 5′ 3′ ylhäältä alas, ja toinen on 5′ 3′ alhaalta ylös.
fyysiseltä kannalta DNA-molekyylit ovat negatiivisesti varautuneita (kaikki nuo fosfaatit), ja normaalisti oikeakätinen helix., Tässä normaali (kutsutaan myös ”B” lihakkuus) valtion, yksi täysi kierre molekyyli käsittää 11 emäsparia, jossa on 0,34 nm: n välillä kunkin nukleotidin pohja. Jokainen typpipitoiset emäkset ovat tasomainen, ja kun pariksi täydentäviä pohja, muodostaa tällä planar ”soitettiin” ”tikkaat” DNA. Ne ovat kohtisuorassa DNA: n pituusakseliin nähden. Suurin osa vapaa-kelluva DNA solun, ja useimmat DNA: ta tahansa vesiliuosta lähellä-fysiologinen osmolariteetin ja pH, löytyy tämä B-konformaatiossa., Muita konformaatioita on kuitenkin löydetty, yleensä hyvin erityisissä ympäristöolosuhteissa. Pakattu lihakkuus, A-DNA: ta, havaittiin artefakti in vitro-kiteytys, hieman enemmän emäkset vuorollaan, puolestaan lyhyempi pituus, ja pohja-paria, jotka eivät ole kohtisuorassa pituusakseliin nähden. Toinen, Z-DNA: ta, näyttää muodossa ohimenevästi GC-rikas DNA: n pätkiä, jotka, kiinnostavaa kyllä, DNA kiertyy vastakkaiseen suuntaan.
on arveltu, että sekä DNA: n A-että Z-muodot ovat itse asiassa fysiologisesti merkityksellisiä., On näyttöä siitä, että A-muoto voi esiintyä RNA-DNA-hybridin kaksoiskierteissä sekä silloin, kun DNA: ta kompleksoidaan joillekin entsyymeille. Z-konformaatio voi tapahtua DNA: n metylaation seurauksena. Lisäksi ”normaali” B-DNA: n rakenne on eräänlainen idealisoitu rakenne perustuu täysin sammutettua, on varmasti hyvin todennäköisesti solun sisällä. Kuitenkin, että nesteytys valtion on jatkuvasti, vaikkakin minuuttiaikataulun, joten DNA: n rakenne on usein hieman vaihdella päässä B-konformaatiossa parametrit Luku \(\PageIndex{2}\).,
prokaryootit, DNA löytyy sytoplasmassa (melko ilmeinen, koska ei ole muita vaihtoehtoja näitä yksinkertaisia organismeja), kun taas aitotumallisilla DNA on löytynyt tuman sisällä. Eroista huolimatta niiden sijainnit, suojan ulkoisia voimia, ja ennen kaikkea, niiden koot, sekä prokaryoottisesta ja eukaryoottisesta DNA on pakattu proteiineja, jotka auttavat järjestämään ja vakauttaa yleinen kromosomin rakenne., Prokaryoottisten kromosomipakkausten suhteen ymmärretään suhteellisen vähän, vaikka joidenkin prokaryoottisten ja eukaryoottisten kromosomien proteiinien välillä on rakenteellisia yhtäläisyyksiä. Siksi useimmat solubiologian kurssit pitävät kiinni eukaryoottisesta kromosomipakkauksesta.

Alasti DNA, onko prokaryoottisten tai eukaryoottisten, on erittäin ohut lohkon materiaali, noin 11 nm halkaisijaltaan. Kun kuitenkin otetaan huomioon eukaryoottisten genomien koko, jos DNA varastoituisi siten tuman sisälle, siitä tulisi hallitsemattoman sekainen. Kuvan ämpäri, johon olet heittänyt sata metriä lankaa ilman mitään aikomusta järjestää sen kelauksessa tai niputtamista se., Nyt harkita, onko sinulla olisi mahdollisuus päästä osaksi ämpäri vetää yksi lohko, ja odottaa vetää vain yksi säie, tai jos sen sijaan olet todennäköisesti vetää ainakin pieni vyyhti lankaa. Solu tekee lähinnä mitä tekisit lanka pitää se järjesti: se on pakattu siististi pienempiin, hallittavissa vyyhtiä. DNA: n tapauksessa jokainen kromosomi kiertyy histonikompleksin ympärille muodostaen kromosomiorganisaation ensimmäisen kertaluvun: nukleosomin.
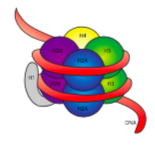
30 nm kuitu on kasassa kaksi sarjaa vuorovaikutusta. Ensinnäkin linker histone, H1, kokoaa nukleosomit yhteen noin 30-nm rakenteeksi., Tämä rakenne on niin vakiintunut disulfidisidoksia, jotka muodostavat välillä H2A histoni yksi nucleosome ja histoni H4 sen naapuri.
histonit ovat perus-(positiivisesti varautuneiden) proteiinien heimo. Ne kaikki toimivat ensisijaisesti järjestää DNA, ja nucleosome muodostuu, kun DNA-huivi (hieman yli 2 kertaa) ytimen ympärille kahdeksan histones – kaksi kunkin H2A, H2B, H3 ja H4. Määrä ja sijainti positiivinen maksut (lähinnä lysines ja arginines) ovat ratkaiseva niiden kyky tiukasti sitoa DNA: ta, joka, kuten aiemmin huomautti, on erittäin negatiivisesti varautuneita., Se, että ”vastakohdat vetävät puoleensa” – idea ei ole vain neuvontapalstojen deittivinkki.

tutkittaessa 3D-rakenne histoni core monimutkainen, me näemme, että kun suhteellisen varaukseton proteiini vuorovaikutus verkkotunnuksia pidä histones yhdessä keskellä, positiivisesti varautuneita jäämiä on löytynyt ympäri ulkopuolella monimutkainen, käytettävissä vuorovaikutuksessa negatiivisesti varautuneita fosfaatit DNA: ta.,
myöhemmässä luvussa keskustellaan siitä, miten entsyymit lukevat DNA: ta siirtääkseen sen tiedot pienempiin, helpommin hallittaviin RNA: n osiin. Toistaiseksi on vain tiedostettava, että kulloinkin suuri osa DNA: sta pakataan tiukasti pois, kun taas osa DNA: sta ei. Koska osat, jotka ovat käytettävissä käyttää voi vaihdella riippuen siitä, mitä on tapahtumassa/solu kerrallaan, pakkaus DNA on dynaaminen. Siinä on oltava mekanismi, jonka avulla voit nopeasti löysää sitova DNA histones, että kun DNA: ta tarvitaan geenien ilmentyminen, ja kiristä sitova, kun se ei ole., Kuten käy ilmi, tämä prosessi liittyy asetylointi ja deasetylointi histonien.

Histoni Acetyltransferases (Hatut) ovat entsyymejä, jotka paikka asetyyli-ryhmä on lysiini on histoni proteiinia. Asetyyliryhmät ovat negatiivisesti varautuneita, ja asetylaatio ei ainoastaan lisää negatiivisesti varautunutta ryhmää, vaan myös poistaa positiivisen varauksen lysiinistä. Tämä on se vaikutus, ei vain neutraloivat pisteen vetovoima välillä, proteiinin ja DNA: n, mutta jopa hieman yllättäen se (kuten maksut)., Toisella puolella mekanismi, Histoni Deactylases (HDACs) ovat entsyymejä, jotka poistavat asetylointi, ja siten palauttaa vuorovaikutusta histoni proteiinia ja DNA: ta. Koska nämä ovat niin tärkeitä entsyymejä, se järkeenkäypää, että he eivät saa toimia tahtoen tai tahtomattaan tahansa käytettävissä histoni, ja itse asiassa, ne ovat usein monimutkaisia muita proteiineja, jotka ohjaavat ja koordinoivat aktivointi muiden prosessien, kuten aktivointi transkriptio.