
Johdanto
Telomeerit ovat nukleoproteiini rakenteita erittäin päät lineaarinen eukaryoottisesta kromosomeja. Ne ratkaisevat samalla kaksi suurta loppuongelmaa. Ensimmäinen koskee kromosomipään suojaa. On arvioitu, että normaalien ihmissolujen on korjattava vähintään 50 endogeenista kaksoissokkoutumista solua kohti solusykliä kohti (Vilenchik ja Knudson, 2003)., Telomeerit erottaa luonnollinen kromosomien päissä haitallisilta DSBs ja estää niiden kohdunulkoinen korjaus, esim., end-to-end-fusions kromosomeja (vanSteensel ja deLange, 1997). Toinen on loppureplikaatio-ongelma, joka käsittelee asianmukaisten telomeerien pituuksien ylläpitoa. Tämän tunnusti itsenäisesti kaksi tutkijaa (Watson, 1972; Olovnikov, 1973). Koska replicative DNA-riippuvaista DNA-polymerases ei voi suorittaa DNA-synteesiä hyvin päät kromosomien, korvaus replicative telomeeri järjestyksessä tappio on tullut RNA-riippuvainen DNA polymeraasi., Tämä entsyymi, nimeltään telomeraasi, yhdessä ensimmäisen telomeeri minisatellite järjestyksessä, löydettiin ripsieläintä Tetrahymena (Blackburn ja Sappea, 1978; Greider ja Blackburn, 1985). Tämä on kuitenkin vain yksi osa telomeeripituuden huolto. Epigeneettinen sääntely telomeeripituuden homeostaasiin, mukaan lukien vuorovaikutus kauan noncoding telomeric toista, joka sisältää RNA ja exonuclease toimintaa polkuja, on myös tutkittu laajasti, koska sen terapeuttinen potentiaali (Wellinger ym., 1996; Polotnianka ym., 1998; Pfeiffer ja Lingner, 2012).,
Telomeraasi on entsyymi, joka vastaa lisäämällä telomeeri toista sekvenssit 3′ loppuun telomeres, on säilytetty monimutkainen entsyymi lukuisia osia . Periaatteessa vain kaksi tärkeimmät komponentit ovat välttämättömiä telomeraasin entsymaattista toimintaa, katalyyttisesti aktiivinen proteiini komponentti, nimeltä telomeeri käänteiskopioijaentsyymin (TERT), ja mallin komponentti, joka muodostuu telomeraasin RNA-alayksikön (TR). Vaikka TERT on evolutionaarisesti melko hyvin säilynyt, TR on hyvin vaihteleva, pituudet vaihtelevat ca. 150 nt (Tetrahymena) yli 2 000 nt (neurospora-suvun sienet)., Vain lyhyt alue koko TR-molekyylissä toimii mallina juuri syntetisoidulle telomere DNA: lle (Greider and Blackburn, 1985; Qi et al., 2013). Tällä alueella TR on yleensä muodostettu täydellinen telomeeri motiivi seurasi osittainen yksi, jälkimmäinen toimii hehkutus alueen nykyisten telomeeri-DNA: ta. Vaikka periaatteessa tarvitaan vain yksi ylimääräinen nukleotidi (osittaisena motiivina), löytyy yleensä useampi kuin yksi. Esimerkiksi kaksi ylimääräistä nukleotidia muodostavat hehkutusaiheen hiirillä tai viisi ihmisellä(Blasco et al., 1995; Feng ym., 1995)., Kasveja, kuitenkin, koko mallin alueella on muuttuja, esim., kaksi Arabidopsis thalianan, seitsemän Arabis sp. tai kuusi nikotianassa (Fajkus et al., 2019). Muilla TR-alueilla on rakenteellisia, sääntelyyn ja proteiineihin liittyviä interaktiivisia toimintoja . Katso myös kaavakuva telomeraasista ja sen aktiivisuusjaksosta kuvassa 1.

Kuva 1 Kaavamainen esitys telomeraasin aktiivisuus sykli kanssa Arabidopsis-tyyppi telomeeri-malli. TERT, Telomeeri Käänteiskopioijaentsyymi; TR, telomeraasi RNA-alayksikkö., Kuva perustuu Sekhriin (2014).
kuinka muuttuvia ovat Telomeerisekvenssit?
Telomeeri-sekvenssit ovat yleensä lyhyitä minisatellites tandemly järjestetty, yleensä seuraavan kaavan mukaisesti (TxAyGz)n. Se minisatellite järjestely on peräisin tapa, joka telomeraasin syntetisoi DNA: ta, lyhyesti sanottuna, ja enimmäkseen samat kuviot, yksi kerrallaan., Useita hypoteeseja, katsovat, että tällainen järjestely on tärkeää, koska se edistää tunnustamista telomeeri erityisiä proteiineja, joita homo – ja heterodimers ja mahdollista muodostaa G-quadruplexes, jotka voivat vakauttaa kromosomin päät tai palvella substraatteina telomeeri-erityiset proteiinit (Spiegel et al., 2020; Tran et al., 2013). Telomere-sekvenssit säilyvät hyvin evoluution kautta, ja suuret eliöryhmät käyttävät ryhmälle tyypillistä telomere-motiivia rakentaakseen telomere-DNA: nsa., Vähitellen kasvava määrä tutkimuksia ja suuret näytökset ovat osoittaneet, että kaikki tutkitut selkärankaiset ja monet pohjapinta metazoans käyttää TTAGGG (Meyne et al., 1989; Traut ym., 2007), kun taas euarthropodalla (niveljalkaiset), myös heksapodalla (hyönteiset), on TTAGG (Frydrychova et al., 2004; Vitkova ym., 2005). Tasaisesti, lukuisia poikkeuksia on kertynyt ajan mittaan, esim., (A(G)1-8) vuonna Dictyostelium (Emery ja Weiner, 1981), TTAGGC vuonna suolinkainen (Nematoda) (Muller et al., 1991), Tcagg in Coleoptera (kovakuoriaiset) (Mravinac et al., 2011), TAGGGG/TAAGG/TAGGGG in Giardia (diplomonads) (Uzlikova et al.,, 2017) tai Ttnnnaggg in Yarrowia clade (hiivat) (Cervenak et al., 2019). Lisäksi, telomeraasi-itsenäistä järjestelmää, jossa minisatellite telomeeri järjestys on kadonnut ja korvattu monimutkainen toistaa, ovat edustettuina, esimerkiksi Hyönteiset ja Uhkaavat (uudelleen (Mason et al., 2016)). Eukaryoottista telomeerisekvenssiä koskevan yleisen tarkastelun osalta ks. (Fajkus et al., 2005; Fulneckova ym., 2013).
Telomeren koostumus kasveissa on vielä monipuolisempi., Tässä käytämme termiä ”kasvit” laajasti ymmärrettynä, joka tunnetaan myös nimellä Archaeplastida tai-britannia Plantae sensu lato, ja käsittää Rhodophyta (punainen levät), Glaucophyta, että Chlorophyte levät luokalla ja Streptophyte levät luokalla (yhteensä nimellä vihreä levä), ja Embryophyta (maa-kasvit) (Tuhat Kasvi Transcriptomes Aloite, 2019). Tyypillinen telomeeri kasvien järjestys on TTTAGGG, jota kutsutaan myös Arabidopsis-tyyppi (tai yksinkertaisesti, kasvi-tyyppi), koska se löydettiin vuonna Arabidopsis thalianan (Richards ja Ausubel, 1988) ja nyt monet muut lajit lähes kaikissa tehtaan tilaukset., Vaikka TTTAGGG on edelleen yleisin, on huomattavaa vaihtelua siten, että telomeeri-sekvenssit punainen ja vihreä levien suvusta. Kuten punalevää (Rhodophyta), telomeeri järjestyksessä tieto on enimmäkseen puuttuu tai hajanaisia, vaikka jotkut telomeeri ehdokkaat on löydetty in silico, kuten AATGGGGGG varten Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T) AGGG galdieria sulfuraria (Fulneckova et al., 2013); TTAGGG on löydetty genomista lukee Porphyra umbilicalis (Fulneckova et al., 2013), mutta tarvitaan lisää todisteita niiden lopullisen aseman vahvistamiseksi kromosomeissa., Telomeren monimuotoisuus viherlevissä heijastaa sekä dynaamisia muutoksia että sen parafyleettistä luonnetta. Vaikka TTTAGGG vallitsee Bcf, kuten sukujen Ostreococcus (Derelle et al., 2006) ja Chlorella (Higashiyama et al., 1995), monet muut erilaiset kuviot on havaittu myös siellä, kuten TTAGGG vuonna suvun Dunaliella ja Stephanosphaeria (Fulneckova et al., 2012) ja Ttttagg in Chlamydomonas (Petracek et al., 1990). Basal Streptophytassa (Klebsormidiofyceae) on kuvattu progressiivisia muutoksia kuvioissa tttaggista TTTTAGGGIIN ja TTTTAGGIIN., Läsnäolo TTAGGG vuonna Rhodophyta ja Glaucophyta johtaa hypoteesi, että tämä on esi motiivi kasvit (Archaeplastida) (Fulneckova et al., 2013).
Koskevat maa kasvit, yksi ensimmäisistä seulonnat suoritetaan osoitti, että Arabidopsis-tyyppinen sekvenssi oli yleisin ja oli enimmäkseen säilytetty läpi niiden fylogenia (Cox et al., 1993; Fuchs ym., 1995), vaikka jotkut näistä tekijöistä oli jo havaittu useita poikkeuksia perheen Amaryllidaceae (entinen Alliaceae), jossa Arabidopsis-tyyppinen sekvenssi oli poissa useita lajeja., Myöhemmin, ensimmäinen telomeeri järjestyksessä epätavallinen maa kasvit, selkärankaiset-tyyppi TTAGGG, oli tunnettu siitä, Aloe ja joissakin muissa Asparagales (Weiss ja Scherthan, 2002; Puizina et al., 2003; Sykorova ym., 2003c). Asparagalesin Tttagg-ja TTAGG-telomere-sekvenssin toistuvista menetyksistä ja takaisinperinnöistä laadittiin hypoteesi (Adams et al., 2001). Kanssa postrefinement, jotta Asparagales vuonna APGIII (Angiosperm Phylogeny Group 2009) (Bremer ym.,, 2009), se oli osoittanut, että vain kaksi suurta evoluution kytkimet telomeeri järjestyksessä koostumus tapahtunut (eikä useita toistuvia voitot ja tappiot), seuraavassa järjestyksessä: ensimmäinen perhe Iridaceae, jossa siirtyminen kasvi-tyyppi TTTAGGG selkärankaisten-tyyppi TTAGGG tapahtui, jonka jälkeen perheet Xeronemataceae, Asphodelaceae ja ydin Asparagales (mukaan lukien Amarillidaceae s.l ja Asparagaceae s.l.,); ja toinen sisällä alaheimoon Allioideae (aiemmin käsitelty erillisenä perheen Alliaceae), jossa täysin uusi telomeeri järjestyksessä syntynyt, CTCGGTTATGGG (Fajkus et al., 2016). Ulkopuolella Asparagales, uusi telomeeri-sekvenssit on havaittu myös maa-kasvi ryhmiä, kuten erilaisia kuin (i), Solanaceae, jossa telomeeri järjestyksessä Cestrum elegans TTTTTTAGGG oli kuvattu (Sykorova et al., 2003a; Sykorova ym., 2003b; Peska ym., 2008; Peska ym.,, 2015) ja (ii) Lentibulariaceae, jossa suvun Genlisea osoitti huomattavaa monimuotoisuutta joitakin lajeja ominaista Arabidopsis-tyyppi telomeeri toistaa, kun taas toiset näytteille sekoittuneet järjestyksessä vaihtoehdot TTCAGG ja TTTCAGG (Tran et al., 2015).
kaikista havaituista telomere motif-poikkeuksista huolimatta maakasvien telomeeristen sekvenssien todellista monimuotoisuutta todennäköisesti aliarvioidaan suuresti. Tuore julkaisu (Vitales et al.,, 2017), joka on seulonta maa kasvi telomeeri sekvenssit tehtiin, todettiin, että telomeeri sekvenssit olivat ainoa tunnettu selvästi alle 10% lajeista ja 40% sukujen sisältämä Kasvi rDNA-tietokanta (www.plantrdnadatabase.com), resurssi tarjoaa molekyyli sytogenetiikka tietoa maakasvit (Garcia et al., 2012). Yhteenveto telomeeri järjestyksessä jakelu kasveja, seuraavat APG IV (Angiosperm Phylogeny Group, 2016) (Byng ym., 2016), sekä tuorein kasvi phylogeny (One Thousand Plant Transcriptomes Initiative, 2019) löytyy kuviosta 2.,
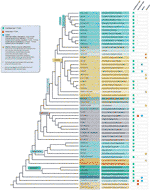
Kuva 2 Telomeeri motiiveja Archaeplastida (kasvit laajassa merkityksessä), joka perustuu APG IV (Angiosperm Phylogeny Group 2016) ja Tuhat Kasvi Transcriptomes Aloite (2019). Haaran pituudet eivät ilmaise reaaliaikaisia asteikkoja., Yksinkertaisuuden ja säästää tilaa, tiettyjä polyphyletic ”ryhmät” (laadut), jotka on merkitty tähdellä puu on ollut edustaa vain yksi haara; samasta syystä, useita pieniä tilauksia (lueteltu sininen neliö vasemmassa yläkulmassa kuva) ovat ei kuvattu puu. Ensimmäinen vihje etiketti tarkoittaa yleensä tehtaan tilaukset ja muutamissa tapauksissa, rajapintojen, laadut ja jopa perheitä, toinen etiketti näyttää edustavia perheitä ja joissakin tapauksissa, edustaja tilauksia tai sukuihin.
näytöksistä Discoveryyn: miten Telomeeriset motiivit voidaan tunnistaa?,
näyttöä siitä, että tietyn ehdokkaan järjestys on todellinen telomeric yksi sisältää useita vaiheita, että oikein julistaa sen lokalisointi kaikkien kromosomien päät, ja lopulta osallistuminen telomeraasien sen synteesiä. Molekyyli sytogenetiikka (lähinnä Fluoresenssi in situ-Hybridisaatio, FISH) on tullut tärkeä visualisointiin terminaali lokalisointi merkitty luotaimet ehdokas sekvenssit ollenkaan kromosomi-termini. Kuitenkin itsenäinen kala se ei riitä todistamaan hyvin terminaalinen kanta., Esimerkiksi, AcepSAT356 oli ehdotettu sipuli telomeeri ehdokas, joka perustuu tuloksiin KALA-analyysi (Pich ja Schubert, 1998). Kuitenkin, sen terminaalinen sijainti in situ ei ole koskaan ollut vakuuttavasti liittyy telomeerien toimintaa. Itse asiassa, löytö Allium minisatellite telomeeri järjestyksessä CTCGGTTATGGG ja telomeraasin tarkoittaisi, että AcepSAT356 on subterminal (Fajkus et al., 2019)., Positiivinen KALA telomeric signaalit voivat myös peittää pieniä muutoksia telomeeri-aiheita, kuten yhden nukleotidin polymorfismeja, tai vääriä negatiivisia tuloksia voivat johtua lyhyet telomeerit on alle määritysrajan tekniikka.
on kaksi muuta lähestymistapaa, jotka määrittävät terminaalin sijainnin suuremmalla resoluutiolla kuin kalat; nämä perustuvat eksonukleaasibal31: n aktiivisuuteen., Ensimmäinen on klassisen Terminal Restriction Fragment (TRF) analyysi, jossa näytteitä käsitellään BAL31 osoittavat asteittainen lyhentäminen terminaali palasia ja vähentää signaalin voimakkuus kasvaa aika exonuclease hoito. Myöhempi analyysi fragmentti pituudet suoritetaan Southern-blot-hybridisaatiolla (Fojtova et al., 2015). Toinen on vertaileva genomin skimmaus (NGS data) nondigested ja BAL31-pilkottu genominen DNA, rinnakkain., Vuonna BAL31 käsitelty aineisto on merkittävä aliedustus telomeeri sekvenssit, siksi terminaali sekvenssit on merkitty verrattuna käsittelemättömän aineisto, käyttäen bioinformatiikan työkaluja RepeatExplorer tai Tandem Toistaa Finder .
tietyn telomeerisekvenssikandidaatin toinen tärkeä testi lajissa on telomeraasin aktiivisuuden osoittaminen. Tässä hyödyllinen kokeellinen lähestymistapa, kehitti ensimmäiset ihmisen solut, on Telomeeri Toista Vahvistus Protocol (TRAP) (Kim et al., 1994), jota seuraa Havaittujen tuotteiden sekvensointi (Peska ym.,, 2015; Fajkus ym., 2016), joka on hieman vähemmän herkkä väärille positiivisille tuloksille kuin kala. Kaikki nämä menetelmät, mukaan lukien kalat (Fuchs et al., 1995; Shibata and Hizume, 2011) ja muut kuten slot-blot hybridisaatio (Sykorova et al., 2003c) ja TRAP (Fulneckova ym., 2012; Fulneckova ym., 2016), voidaan käyttää näytön telomeerit koko laaja ryhmät monimutkaisia organismeja, kuten kasveja., Kuitenkin, vain yhdistelmä sopivasti valittu menetelmät voivat johtaa vakuuttavasti johtopäätös telomeeri-toiminto ehdokas järjestyksessä, koska tulokset pohjan yksi lähestymistapa voisi olla harhaanjohtava. Täydellisempi katsaus strategioita de novo telomeeri ehdokas sekvenssin tunnistaminen, mukaan lukien ensimmäinen yritys Tetrahymena (Greider ja Blackburn, 1985) on esitetty metodologinen artikkeli, jossa painotetaan NGS lähestymistapa käyttää kasveja, joilla on erittäin suuri genomit (Peska et al., 2017).
onko Telomere-jaksoissa Homoplasia?,
esi-isien telomeerisekvenssin arvellaan olevan TTAGGG, ja se esiintyy yleisimmin elämän puun (Fulneckova et al., 2013). Silti näyttää selvältä, että homoplasman taajuus telomere motif-evoluutiossa on suhteellisen korkea. Esimerkiksi, lyhyitä, yksinkertaisia kuvioita, kuten kasvi-tyyppi TTTAGGG on ilmestynyt itsenäisesti ja toistuvasti cryptomonads, oomycete sienet, ja alveolates; samoin selkärankaisten-tyyppi TTAGGG on syntynyt toissijaisesti tiettyjen ryhmien kasveja (Asparagales, Rodophyta ja Bcf levät) (Sykorova et al., 2003c; Fulneckova ym.,, 2012; Fulneckova ym., 2013; Somanathan ja Baysdorfer, 2018). Syy jotkut telomeeri-sekvenssit on syntynyt useammin kuin muut, yleensä monimutkaisempia sekvenssejä liittyy luultavasti valinta paineet, mikä olisi hyväksi tarkkuus tietty sekvenssi-spesifisiä DNA-proteiini vuorovaikutus (Forstemann et al., 2003). Jos jokainen telomeerimotiivi muuttuisi, telomeerisen nukleoproteiinirakenteen häiriöt johtaisivat väistämättä genomin epävakauteen., Tämä on syy, telomeeri-sekvenssit ovat niin evoluution vakaa, sisältää hyvin vähän romaani ja onnistuneen sekvenssit, kuvio mukainen ajatus toistuvat tappiot ja syntyy tyypillinen telomeeri sekvenssit, kuten ehdotettu Asparagales (Adams et al., 2001).
löytäminen homoplasy koko telomeeri sekvenssit herättää kysymyksen, mitkä ovat molekyyli-syyt ja prosessit tapahtuvat aikana nämä muutokset? Muutos telomeren sekvenssissä, vaikka se joissakin tapauksissa tuntuu vähäpätöiseltä (esim., yksi ylimääräinen T), voi aiheuttaa vakavia häiriöitä genomin eheyttä, koska häiriintynyt tasapaino telomeeri-DNA-proteiini-vuorovaikutuksia. On myös epäselvää, voiko telomeerisarjan muutoksella olla evolutionaarista etua; tässä suhteessa (Tran et al., 2015) ehdotti, että ulkonäkö ”methylatable” sytosiini G-rikas telomeeri strand olisi nostaa mahdollisuus asetuksen epigeneettisiä muutoksia.
mitkä ovat Telomeren motiivien muutosten Molekyylisyyt?,
telomeerisekvenssimuutoksen selittämiseksi ensimmäinen ehdokas on telomeraasin, telomeraasi RNA: n (TR) mallialayksikkö. Aiemmin havaitut TR hiivasta ja selkärankaisten kuuluu eri ryhmään selostukset, joiden yhdistävä piirre oli, että he olivat transkriptoidaan RNA-polymeraasi II (Pol II)—in kaikki, mutta ripsieläimiä; tämä oli yksittäinen poikkeus Pol II selostukset ennen julkaisemista maa kasvi TR tunnistaminen . Käyttämällä suhteellisen pitkä telomeeri motiivi Allium etsiä sen TR sisällä koko RNA-sekvenssin tiedot-allas, Fajkus et al., (2019) osoitti, että aiemmin tunnettu noncoding RNA mukana stressi reaktio A. thalianan, nimeltään AtR8, oli todellakin telomeraasin RNA-alayksikön (Wu et al., 2012; Fajkus et al., 2019). Se oli RNA polymeraasi III: n (Pol III) transkriptio, joka sisälsi sen promoottorirakenteen vastaavat säätelyelementit. Pitkään, tutkijat odotettavissa, että kasvi TR olisi niin erilaiset, että se olisi mahdotonta tunnistaa se perustuu homology haku (Cifuentes-Rojas et al., 2011)., Jonkin verran samankaltaisuutta käytettiin kuitenkin onnistuneesti useiden Alliumlajien yhteisen TR: n tunnistamiseen vertailevalla Blastilla. Yllättäen järjestyksessä homologia, läsnäolo saman sääntelyn elementtejä, ja vastaava malli led-alueen tunnistaminen TRs vuonna Allium, Arabidopsis ja yli 70 muuta kaukaista sukua kasveja, mukaan lukien ne, joilla on eriytynyt telomeeri-aiheita, kuten Genlisea, Cestrum, ja Tulbaghia., Tietääksemme levästä TR ei ole vielä tietoa, mikä selvittäisi, onko Pol III TR-transkriptio yleinen ominaisuus kaikille kasveille vai ei. This work (fajkus et al., 2019), joka perustuu CRISPR knock-out ja muut kokeet, myös osoitti, että aiemmin havaitut telomeraasin RNA-ehdokas A. thalianan (Cifuentes-Rojas et al., 2011; Beilstein ym., 2012) ei ollut telomeraasin toiminnallinen mallialayksikkö, kuten myös pian sen jälkeen (Dew-Budd et al., 2019)., Olettaen, että Pol II/Pol III riippuvuus TR transkriptio on luotettava evoluution merkki, tulevaisuuden TR tutkimuksen muut keskeiset aitotumallisilla suvusta todennäköisesti avata uusia oivalluksia alkuperän eukaryooteissa. Telomeraasi geenit ja telomeeri-sekvenssit ovat tunnistamattomia tietolähteitä tähän suuntaan, ja löytää Pol III riippuvainen TR biogenesis koulutusjakson ripsieläintä ja kasvin suvusta voi edustaa ensimmäiset askeleet tähän suuntaan (Greider ja Blackburn, 1989; Fajkus et al., 2019).
miten kromosomit muuttuivat lineaarisiksi?,
valtaosa prokaryooteista sisältää kiertokromosomeja, kun taas lineaarikromosomit ovat sääntö eukaryooteissa., Näin ollen on olemassa kaksi mahdollista skenaariota, joissa joko (i) linearisointi oli suorittaa primitiivinen telomeraasi, ennen muita prosesseja, jotka johtivat nykyisen lineaarinen kromosomien ominaisuuksia ja toimintoja, tai (ii) linearisointi ennalta aitotumallisilla pyöreä kromosomi oli aluksi telomeraasin itsenäinen, mutta juuri ennen kuin nykyinen eukaryooteissa eriytynyt, primitiivinen telomeraasin alkoi miehittää kromosomin päät ja tuli olennainen äskettäin muodostettu lineaarinen kromosomien (Nosek et al., 2006). Villasante ym., (2007) ehdotti evoluution skenaario, jossa rikkoutuminen esi prokaryoottiset pyöreä kromosomi aktivoitu osaksi mekanismi DNA-päät, jolloin muodostumista telomeres, joita rekombinaatio-riippuvainen replikoinnin mekanismia: seuraukset tämä hypoteesi johtanut yllättävää tulokseen, että aitotumallisilla sentromeerien olivat peräisin telomeres.
Mielenkiintoista, vastapäätä prosessin linearisointi, eli muodostumista pyöreä kromosomit (kutsutaan myös rengas kromosomit) on syntynyt ajoittain aikana kehitys eukaryooteissa, vaikka on erittäin epävakaa., Esimerkiksi, jos Amaranthus tuberculatus, rengas kromosomit esiintyi stressin aiheuttama vaste, kuljettaa vastustuskyky rikkakasvien (glyfosaatti); nämä ylimääräiset rengas kromosomit eivät näytä hybridisaatio telomeeri-antureita karyotyypin analyysi (Koo ym., 2018). Lähes universaali telomeraasi-järjestelmä ja erilainen kuin muut pyöreä aitotumallisilla kromosomien eivät salli meidän tuki yksi hypoteesi yli muiden., Kuitenkin recombinational koneita käytetään vaihtoehtoisten pidentäminen telomeerit (ALT), on telomeraasi-riippumaton polku, joka liittyy tiettyjen ihmisten syöpiin (Zhang et al., 2019), esiintyy jo prokaryotesissa. Lisäksi on näyttöä siitä, kromosomi linearisointi tapahtuu itsenäisesti eri prokaryote suvusta (Ferdows ja Barbour, 1989; Nosek et al., 1995; Volff ja Altenbuchner, 2000). Näin ollen hypoteesi, että ensimmäinen lineaarinen aitotumallisilla-kromosomi (peräisin prokaryote esi-isä) oli telomeraasi-riippumaton näyttää todennäköisemmältä., On joitakin esimerkkejä, jotka osoittavat, että telomeraasi-pohjainen järjestelmä ei ole välttämätöntä, että telomeeri ylläpito kaikilla aitotumallisilla: retrotransposons Drosophila telomeres, satelliitti toistaa Chironomus, toinen hyönteinen (Rubin, 1978; Biessmann ja Mason, 2003), ja ALT telomeraasi-negatiivinen ihmisen syövistä (Hu et al., 2016; Zhang ym., 2019). Vielä, jotkut nämä järjestelmät eivät ehkä ole niin erilaisia, ja ehkä on yhteinen alkuperä: Drosophila, telomeeri ylläpito, joka perustuu retrotransposition, ei ole liian erillään telomeraasi-pohjainen mekanismi (Danilevskaya et al.,, 1998), joka johti olettamukseen, että telomeraasi itsessään voi olla entinen retrotransposoni. Mutta varmasti, telomeraasi-negatiivinen kasvilajeja ei ole havaittu tähän mennessä ja kaikki poikkeukset, jossa tyypillinen kasvi-tyyppi telomeeri oli poissa, olivat myöhemmin osoittanut, että on erilainen, mutta silti telomeraasi-syntetisoitu, kuviot. ALT-koneita on kuitenkin laitoksissa telomeraasin toiminnan rinnalla (Watson and Shippen, 2007; Ruckova et al., 2008). Mielenkiintoisia kysymyksiä rooli telomeraasin, telomeerit ja niiden huolto kasvi kasvaimet johtuvat siitä., Houkutteleva yksi on noin puuttuminen etäpesäke kasveissa, vaikka läsnäolo ALT, ehkä liittyvät kasvin kudoksen jäykkyyttä tai eri immuunijärjestelmä kuin eläimillä (Seyfried ja Huysentruyt, 2013).
vaikka saamme yhä enemmän tietoa telomeeribiologiasta, emme silti pysty selittämään telomeraasin syntyä eukaryooteissa., Nykyinen näyttö tukee oletusta, että syntyminen eukaryooteissa yhdessä niiden lineaarinen kromosomien, telomeres ja telomeraasien liittyi ulkonäkö spliceosomal intronit vuonna archaeal isännät (Koonin, 2006; Fajkus et al., 2019). Tertin ja muiden retroelementtien samankaltaisuudesta on keskusteltu jo jonkin aikaa(Pardue et al., 1997). Merkillistä, suhteellisen tuore tutkimus osoitti, että TERT, koska todennäköinen jäsen jälkeläisryhmä II intronit, on peräkkäin lähellä Penelope-kuten elementti retrotransposons (Gladyshev ja Arkhipova, 2007)., Mutta TERT on vain yksi kaksi olennaista telomeraasin osia, ja TR on, sen alkuperä, jopa enemmän arvoituksellinen, koska sen alhainen sekvenssikonservoinnista kaikilla aitotumallisilla .
Johtopäätös
alussa kasvi genomiikan aikakaudella, telomeeri järjestys pidettiin lähes muuttumaton. Telomeerien yleinen säilyttäminen ja telomeraasijärjestelmä viittasivat siihen, että kaikilla kasveilla voi olla TTTAGG-kasvityyppinen telomeeri. Tunnistaminen epätavallinen telomeeri-sekvenssit monimutkainen kasvien genomit, monissa tapauksissa jättimäiset C-arvot (kuten Cestrum ja Allium sp.,), oli vaivan arvoista, sillä poikkeuksellisen pitkä Allium telomeeri motiivi oli vihje etsivät aitoa TR maan kasveja. Äskettäin kuvattu TR kasveja ja edelleen telomeeri/telomeraasin tutkimus pohjapinta clades levien saattaa paljastaa arvokasta tietoa siitä, varhainen kehitys, siis kasvi telomeeri tutkimus voi merkittävästi edistää hypoteeseja syntymistä eukaryooteissa.
Kirjoittaja Maksut
VP ja TP ovat vaikuttaneet yhtä lailla kirjallisesti, editointi ja valmistelu tämä mini-review.,
Rahoitus
eturistiriita
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
Kiitokset
Tuhat Kasvi Transcriptomes Aloite (2019). Tuhat kasvitranskriptomia ja viherkasvien fylogenomiikkaa. Luonto 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar