
Johdanto
Sairaalainfektioiden aiheuttama monelle resistentit Acinetobacter baumannii ovat yleistymässä maailmanlaajuisesti, erityisesti tehohoidossa asetus (Wieland et al., 2018). Menestys tämä bakteeri on helpottanut sen kyky selviytyä eri ympäristöissä vaikeuttaa sen nopea kyky hankkia monilääkeresistenssi. Pintahiilihydraatit ovat avainrooleissa A: n yleiskunnossa ja virulenssissa., baumannii (Lees-Miller ym., 2013; Geisinger and Isberg, 2015; Weber et al., 2016). A. baumannii tuottaa korkean molekyylipainon kapselipolysakkaridia (CPS), joka ympäröi ulompi kalvo (Kuva 1) (Russo et al., 2010). Koostuu tiiviisti pakattu toistuva oligosaccharide alayksikön (K-yksikköä), CPS muodostaa erillinen kerros bakteeri-pinta, joka tarjoaa suojan erilaisia ympäristöolosuhteita, auttaa veropetosten isännän immuunipuolustusta ja lisäämään vastustuskykyä useita antimikrobisia yhdisteitä (Russo et al., 2010; Iwashkiw ym., 2012; Geisinger ja Isberg, 2015).,
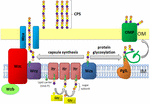
Kuva 1. Kaavamainen esitys kapselin polysakkaridi kokoonpano ja vienti A. baumannii. Synteesi alkaa alkuperäisen transferaasin (Itr; viininpunainen), joka sijaitsee sisemmän kalvon (IM) siirtää ensimmäisen sokeria toistuva yksikkö rasva harjoittaja (Und-S; vihreä ympyrä). Tämän jälkeen kasvavaan yksikköön lisätään sokereita spesifisillä glykosyylitransferaasilla (Gtr; keltainen) sisäkalvon sytosolisella puolella., Kapselin alayksikkö (K-yksikkö) siirtyy tämän jälkeen periplasmaan integraalikalvoproteiini Wzx: n (Tummansininen) kautta. Sokeri alayksikön polymeroidaan, jonka Wzy proteiinia (violetti) ja Wza/Wzb/Wzc monimutkainen (syaani, lime, punainen) koordinaatit korkean tason polymerointi ja viennin kasvava ketju, siirtämällä niitä ulompi kalvo (OM). Glykosylaatiota varten pgll (oranssi) yhdistää k-yksiköt valikoituihin uloimpiin kalvoproteiineihin (OMP; tummanvihreä).
A. baumannii, kapseli kokoonpano ja vienti tapahtuu kautta Wzy-riippuvainen reitti (Hu et al.,, 2013; Kenyon and Hall, 2013; Willis and Whitfield, 2013; Woodward and Naismith, 2016) (kuva 1). Tyypillisesti koostuu 4-6 sokereita, K yksikkö on koottu rasva-operaattorin molekyyli undecaprenyl pyrofosfaatti (Und-S), joka tarjoaa rakennustelineiden kasvava sokeria ketju (Whitfield, 2006). Ensimmäisen sokeria K-yksikkö on palkannut sisempi kalvo (IM)-sidottu alkuperäisen transferaasin (Itr), jonka jälkeen juokseva lisäämällä sokeria kasvava K-yksikkö erityisiä glykosyyli-glukuronyylitransferaasin (Gtr) entsyymit (Kuva 1) (Woodward ja Naismith, 2016)., Jokainen K-yksikkö siirretään sitten periplasmic puolella CHAT, jonka Wzx translocase ja polymeroidaan, jonka Wzy, joka siirtää kasvava polysakkaridi ketju yhdestä Und-S harjoittaja seuraava saapuva alayksikkö (Kuva 1) (Collins et al., 2007). Kun CPS polymeeri on syntetisoitu, se kuljetetaan solun pinnalle kautta hyvin koordinoitu prosessi, johon liittyy vuorovaikutusta kolme proteiineja; Wza, Wzb, ja Wzc, jotka muodostavat vienti-laitteet (Kuva 1)., CPS synteesi on yksi käsi kaksihaarainen polku, koska nämä K-yksiköt ovat myös koristella tiettyjä pinta-proteiinien välityksellä O-liittyvät proteiinien glykosylaatio (Lees-Miller et al., 2013). Tässä tapauksessa, yksi K-yksiköt siirretään vastaanottajan proteiinien O-oligosaccharyltransferase PglL (Kuva 1) (Iwashkiw et al., 2012). A. baumannii, proteiinien glykosylaatio edistää biofilmin muodostumisen parantamalla alkuperäistä kiinnitystä ja kypsymisen biokalvot, ja patogeenisuus kuten on osoitettu useissa eläinten infektio-mallit (Iwashkiw et al., 2012; Scott ym., 2014; Harding ym.,, 2015). Biokalvo on kasvun tila, jossa bakteeri-yhteisöt ovat suljettu sisällä exopolysaccharide matriisi ja sen on osoitettu olevan merkittävä rooli A. baumannii pysyvyys ja kestävyys.
Muut pinta hiilihydraatteja tiedetään vaikuttavan patogeenisuus A. baumannii ovat lipooligosaccharide (LOS) ja exopolysaccharide poly-β-(1-6)-N-acetylglucosamine (PNAG) (Preston et al., 1996; Weber ym., 2016). PNAG muodostaa biofilmien yhtenäisen ”liiman”ja muodostaa huomattavan osan biofilmeistä (Choi et al., 2009; Longo ym., 2014)., Toisin kuin useimmat Gram-negatiiviset bakteerit, A. baumannii ei tuota perinteistä lipopolysakkaridi, mutta sen sijaan samanlainen pinta glyco-konjugaatti, LOS, joka koostuu rasva ydin, joka ei ole O-antigeeni (Kenyon ja Hall, 2013; Kenyon ym., 2014b). LOS-tuotannon menetys A. baumannii: ssa vähentää ulomman kalvon vakautta, mikä heikentää kuntoa (Moffatt ym., 2010; Beceiro et al., 2014).
vaikka monet hiilihydraattimoottorit vaikuttavat patogeenisuuteen, voidaan väittää, että CPS on A. baumannii: n hallitseva virulenssitekijä. Uudelleentarkastelun tarkoituksena on vahvistaa sitä, mitä A: sta tiedetään., baumannii kapseli, mukaan lukien valitut rakenteet, biosynteesi ja gene organisaatio, rooli CPS vuonna virulenssi, ja mahdollisuudet CPS tavoitteeksi tulevalle rokotteiden ja lääkkeiden kehittämiseen.
Geneettinen Organisaatio K Loci
täydellinen genomien enemmän A. baumannii-isolaatit ovat käytettävissä, todellinen monimuotoisuus kapseli rakenteita läsnä sisällä tämä bakteeri on käymässä ilmeiseksi. Tähän mennessä A. baumannii (Kuva 2a) (Shashkov et al., 2017). Nämä alueet ovat tyypillisesti kooltaan 20-35 kb., Analyysi geenejä ohjaamalla CPS synteesi kymmenen täydellinen A. baumannii genomien alun perin johti nimeäminen yhdeksän kapseli tyypit, KL1–KL9, josta tuli perusta universaali kirjoittamalla järjestelmä näistä loci (Kenyon ja Hall, 2013). Järjestelmä on sittemmin laajentunut vastaamaan uuden K loci: n tunnistamista. Kromosomi sijainti K-lokus, välillä fkpA ja lldP geenejä, on erittäin säilytetty välillä A. baumannii-kantoja, ja se sisältää ne geenit tarvitaan biosynteesiin ja vienti kunkin CPS-tyyppi (Kuva 2) (Hu et al.,, 2013; Kenyon and Hall, 2013). Poikkeuksena tähän sääntöön ovat A. baumannii-kantoja, joilla KL19 ja KL39 alueilla, joissa geeni koodaa Wzy-polymeraasi, wzy, löytyy pieni geneettinen saari muualla kromosomi (Kenyon ym., 2016a). Lisäksi CPS: ssä esiintyviin yleisiin sokereihin tarvittavia geenejä löytyy muualta., Kaikki K loci ssa samanlainen geneettinen kokoonpano, erittäin vaihteleva klusterin synteesi ja koodaavan geenien tarvitaan biosynteesiä ainutlaatuinen KL-tyyppi monimutkaisia sokereita, reunustaa toisella puolella on hyvin säilynyt CPS viedä geenejä, ja toisella puolella joukko geenit koodaavat konservoituneita yksinkertaisia sokereita ja lähtöaineiden (Kuva 2A)., Se wzx ja wzy geenejä tarvitaan toista-yksikkö käsittely on erittäin vaihteleva, välillä K loci (Kuva 2A, vaaleansininen), mikä osoittaa, spesifisyys erityisesti K-yksikön rakenteet ja, yleensä, jotta gtr taustatekijät, koodaus erityisiä glycosyltransferases kuluessa KL alueilla, kääntäen vastaa järjestyksessä.
muuttujan alueella joitakin KL gene clusters, kuten KL37 ja KL14, ettei geenien monimutkainen sokeri synteesi, koska ne sisältävät vain yksinkertaisia sokereita niiden K-yksiköt (Arbatsky et al., 2015; Kenyon ym.,, 2015). Moninaisuuttaan lisää myös se, että useat KL-alueet sisältävät tarpeettomia geenejä, joita ei tarvita lopullisen k-yksikön synteesissä. Esimerkiksi KL8 ja KL9 sisältää kaksi itr-geenit (Kenyon ja Hall, 2013) ja KL37 on pgt1 phosphoglyceroltransferase, mutta ei vastaavaa phosphoglycerol jäännös määritetään rakenne (Arbatsky et al., 2015). Lisäksi KL93, kaksi lisäys järjestyksessä elementit (ISAba26 ja ISAba22) keskeyttää pgt1 tekijä (Kuva 2) (Kasimova et al., 2017)., Geenit tarvitaan erityisiä sokeria biosynteesi ei keskustella, koska nämä reitit on käsitelty aiemmin ja eivät kuulu tämän tarkastelun (Hu et al., 2013; Kenyon and Hall, 2013). Lisäksi, muut geenit sijaitsevat KL koodata tuotteita ennustettu olla mukana asetylointi tai acylation muuttaminen erityisiä glycans (Kuva 2A, vaaleanpunainen). Vaikka tarkastelu K loci voi paljastaa paljon K-yksikön rakenteet, kemiallinen analyysi ja biokemialliset testaus on tarpeen määrittää tarkka rakenteita ja tunnistaa erityisiä yhteyksiä toista sokerit.,
CPS Rakenteet
varhaisimmat tutkimukset CPS-serotyypin A. baumannii ajettiin tarve kehittää menetelmä syrjiä A. baumannii eristää muista Acinetobacter-lajit, kuten fenotyyppinen analysointi oli rasiteta epäselvyyttä ja virheellisen tunnistuksen (Traub, 1989). Kuten edellä on kuvattu, on ilmiömäinen monimuotoisuus nähdään A. baumannii CPS biosynteesi gene clusters, mikä merkitsee, että monimuotoisuus nähdään K-yksikkö rakenne (Hu et al., 2013; Kenyon and Hall, 2013). Yhteensä yli 40 erilaista A., baumannii k-yksikkörakenteita on toistaiseksi selvitetty NMR-spektroskopialla ja kemiallisella analyysillä. K-yksikkörakenteissa on eroja sokerin koostumuksessa. Niihin voi kuulua tavallisen UDP-sidossokerin johdannaisia, kuten glukoosia, galaktoosia ja glukuronihappoa, tai harvinaisia ja epätyypillisiä sokereita, kuten ei-2-ulosonihappoja. Rakenteet vaihtelevat pituudeltaan ja voivat koostua vain kahdesta jäännöksestä, kuten K53-tyyppisessä CPS: ssä (Shashkov et al., 2018), tai jopa viisi tai kuusi monosakkarideja, kuten että nähdä K37 (Kuva 2B) (Arbatsky et al., 2015)., Rakenteet poikkeavat toisistaan myös yhteyksiä sekä sisällä ja välillä K-yksiköitä, jolloin tuotanto K-yksiköitä, jotka ovat lineaarisia tai liity sivuoksat, kuten nähdään K1 ja K93, vastaavasti (Kuva 2B) (Kenyon ym., 2016a; Kasimova ym., 2017). Erot sijainti erityisiä glykosidisidoksella joukkovelkakirjoja ja O-asetylointi kuvioita eri oligosakkarideja sisällä rakenne osaltaan myös K-yksikkö monimuotoisuutta.,
Vaihtelu K-yksikön rakenteet voivat olla hiuksenhieno, esimerkiksi, K12 ja K13 eroavat vain sidos kahden glycans, joka edellyttää vaihtoehtoisen Wzy-polymeraasi; näin ollen K-lokuksen molemmat kannat ovat identtisiä lukuun ottamatta wzy-geenin (Kuva 2). Vaihtoehtoisesti vaihtelu voi olla silmiinpistävää, kuten sisällyttäminen harvinaisia sokereita, kuten pseudaminic, legionaminic, tai acinetaminic happojohdannaisten nähty K2/6, K49, ja K12/13 rakenteita, vastaavasti (Kuva 2) (Kenyon ym., 2014a, 2015; Vinogradov et al., 2014; Kasimova ym., 2018)., Mielenkiintoista on, että akinetamiinihappojohdannaiset on tunnistettu vain A. baumannii: ssa, eikä niitä tavata missään muualla luonnossa (Kenyon et al., 2017). Lisäksi, jotkut K-yksiköt sisältävät ainutlaatuinen johdannaiset erityisiä glycans, esimerkiksi pseudaminic happo K93 on asetyloitu kanssa (R)-3-hydroxybutanoyl ryhmä (Kasimova et al., 2017), kun taas K2: ssa ja K6: ssa pseudamiinihappo ei ole asetyloitu (kuva 2)., Lisäksi rakenne K4 on ainutlaatuinen, koska se sisältää vain aminosugars, D-QuipNAc, ja yksi terminaali, N-asetyyli-D-galactosamine (D-GalpNAcA) haara, joka on rajattu kanssa pyruvyl ryhmä, harvinainen aihe ja ensimmäinen kuvattu Acinetobacter (Kuva 2A, musta) (Kenyon ym., 2016b). Kuten määrä K-yksikön rakenteet selvitetty lisää, niin ei luottamusta päätellä, K-yksikön rakenteen analyysi biosynteesin klustereita sijoitettu A. baumannii KL., Kuitenkin, vaikka informatiivinen, ymmärrystä rooli CPS pelaa synnyssä on tärkeää soveltaa tätä tietoa parantaa tuloksia A. baumannii infektiot.
Rooli Virulenssi, Mikrobilääkeresistenssin ja Pysyvyys
ei ole epäilystäkään Siitä, että läsnäolo CPS on välttämätöntä A. baumannii patogeenisuus. Se ei ole vain välttämätöntä isännän immuunipuolustuksen (Russo et al., 2010; Geisinger ja Isberg, 2015), mutta se on tärkeää, että kestävyys antimikrobisia yhdisteitä ja selviytyminen vaikeissa ympäristöissä (Luukas et al., 2010; Russo et al.,, 2010; Geisinger ja Isberg, 2015). CPS välittää immuunijärjestelmän veronkierron monissa A. baumannii-kantoja rajoittamalla vuorovaikutusta immunogeeninen pintarakenteet bakteerit ja isännän puolustusta (Preston et al., 1996; Wu ym., 2009; Russo et al., 2010; Umland ym., 2012; Lees-Miller ym., 2013; Geisinger and Isberg, 2015; Wang-Lin et al., 2017). Poistaminen kapselin useita erilaisia A. baumannii-kantoja on osoittanut, vähentää selviytymisen ihmisen seerumin ja askites nesteen, ja vaimennus rotan ja hiiren infektio mallit (Russo et al., 2010; Umland ym., 2012; Lees-Miller ym.,, 2013; Sanchez-Larrayoz ym., 2017). Lisäksi up-asetuksen kapseli tuotanto yleisesti käytetä A. baumannii kanta ATCC 17978 (K3 CPS type) seerumin resistenssin ja virulenssin hiiren infektio malli (Geisinger ja Isberg, 2015). Lisäksi romaani antimikrobisia hoitoja voitaisiin kehittää erityisiä CPS tyypit, esimerkiksi ne, jotka sisältävät pseudaminic happoa, koska sen läsnäolo on ollut korreloi parannettu virulenssi (Hitchen et al., 2010; Kao et al., 2016).
paitsi suojaa isäntäpuolustukselta, A., baumannii CPS: n tuotanto kasvaa vastustuskykyä erilaisia antimikrobisia yhdisteitä, mukaan lukien ne, joita käytetään desinfiointiin kliinisissä asetukset (Geisinger ja Isberg, 2015; Tipton ym., 2015; Chen et al., 2017). Lisäksi A. baumannii: n kasvu mikrobilääkkeiden inhibitorisissa pitoisuuksissa vaikuttaa CPS: n tuotantoon. Esimerkiksi, altistuminen antibiooteille kloramfenikoli tai erytromysiini johti parannettu kapseli synteesiä ATCC 17978 (Geisinger ja Isberg, 2015) meropeneemi altistuminen valittu mutaatioita, jotka johtavat menetys CPS tuotannon eristää 37662 (Chen et al., 2017)., Tutkimukset suoritettiin laajempi valikoima A. baumannii-kantoja tarvitaan tunnistaa, jos suojan mikrobilääkkeiden tarjoamia CPS on kantakohtainen, kapseli-alustyyppikohtainen, tai universal.
A. baumannii-valmisteen kyky säilyä kliinisessä ympäristössä on epäilemättä lisännyt kolonisaatiota ja infektioita herkillä potilailla. A. baumannii pystyy elossa kuukausia sairaalassa pinnoille, kuten sänky kiskot, huonekalut ja lääkinnällisiä laitteita, jotka tarjoavat säiliö, joka on usein lähde siirto-ja infektio (Wendt et al., 1997; Gayoso ym., 2013)., Merkittävä kuivumisen toleranssi A. baumannii on ajateltu johtuvan ”rintakuva tai boom” – strategia, jossa jatkuva tekemät solujen hengissä kustannuksella kuolevan solut; CPS parantaa kuivumisen suvaitsevaisuutta tarjoamalla fyysinen este, helpottaa veden säilyttäminen (Roberts, 1996; Webster et al., 2000; Gayoso ym., 2013; Bravo et al., 2016). CPS: n suora rooli kuivausresistenssissä osoitettiin äskettäin A. baumannii-kannassa AB5075 (K25 CPS-tyyppi). Tässä tutkimuksessa ab5075: n acapsular-muunnoksessa oli 2.,5-kertainen elinkelpoisuuden väheneminen verrattuna vanhempien kantaan (Tipton et al., 2018). Lisäksi kaksi lähisukulaisiin A. baumannii, Acinetobacter calcoaceticus ja Acinetobacter baylyi, tuotanto exopolysaccharide ja/tai CPS on osoitettu edistää kuivumista survival (Roberson ja Firestone, 1992; Ophir ja Gutnick, 1994). Lisäksi vaikuttaa vastustuskykyä kuivumisesta, CPS on liittynyt muita virulenssi ominaisuuksia, kuten liikkuvuutta (McQueary et al., 2012; Huang et al., 2014) ja biofilmin (Umland et al., 2012; Lees-Miller ym.,, 2013), mikä sementoi sen aseman patogeenisena tekijänä.
Viimeaikaiset tutkimukset ovat sidoksissa vaihe-muuttuja fenotyyppi A. baumannii AB5075 muutoksia CPS: n tuotannon, kuin erittäin virulentti läpinäkymätön vaihtoehtoja tuottaa CPS kerros kaksi kertaa paksuus heidän läpikuultava kollegansa (Chin et al., 2018). Tämä siirtyminen läpikuultava läpinäkymätön myös kasvanut dramaattisesti patogeenisten potentiaalia A. baumannii AB5075. Myös resistenssi tavallisille sairaalan desinfiointiaineille ja aminoglykosidiantibiooteille lisääntyi (Tipton et al., 2015; Chin et al.,, 2018) ja läpinäkymätön vaihtoehtoja olivat myös enemmän vastustuskykyisiä ihmisen lysotsyymi, että cathelicidin-liittyvät antimikrobinen peptidi LL37 ja vetyperoksidia verrattuna läpikuultava siirtomaita (Chin et al., 2018). Lisäksi, läpinäkymätön isolaateista oli lisääntynyt toleranssi kuivunutta ehtoja ja kilpaili läpikuultava kollegansa hiiri infektio malli (Chin et al., 2018). Useita tekijät ovat mukana vaiheen vaihtelu, edelleen tutkimukset on tehty, voit määrittää osuus CPS tuotannon virulentimpaan läpinäkymätön fenotyyppi., Vuonna seuraava julkaisu, kirjoittajat osoitti, että acapsular variantti oli huomattavasti alttiimpia lysotsyymi-ja desinfiointiaineet verrattuna sen läpinäkymätön AB5075 villin tyypin vanhempi (Tipton ym., 2018). Mielenkiintoista, ei ollut eroa vastustuskykyä LL-37 ja vetyperoksidin välillä läpinäkymätön villi-tyyppi ja acapsular kantoja, mikä viittaa siihen, muista tekijöistä kuin CPS: n tuotanto johtaa tämän fenotyypin läpinäkymätön variantteja A. baumannii (Tipton ym., 2018).,
Asetus CPS Tuotannon
Ympäristön vihjeet, kuten lämpötila, osmoottinen paine ja muutokset metaboliitti ja ion saatavuus voi vaikuttaa bakteeri-CPS: n tuotannon (Hagiwara ym., 2003; Lai ym., 2003; Mouslim ym., 2004; Willenborg ym., 2011). Ei ole yllättävää, että muutama sääntelyn mekanismeja on tunnistettu CPS: n tuotannon, kuin CPS-tasot ovat yleisesti säännelty post-translationally kautta fosforylaation CPS vienti koneet (Whitfield ja Paiment, 2003; Chiang et al., 2017). A., baumannii, vain kaksi sääntelyviranomaisten kapseli tuotanto on toistaiseksi tunnistettu; BfmRS ja OmpR-EnvZ, sekä kaksi-komponentti-signaalin transduktio järjestelmiä, jotka pelata useita sääntelyn roolit mukana kirjekuori biogenesis (Geisinger ja Isberg, 2015; Tipton ja Melko, 2017; Geisinger et al., 2018). Kun siihen kohdistuu antibiootti paine, A. baumannii ATCC 17978 cps: n ilmentyminen oli lisääntynyt, BfmRS-riippuvaisella tavalla (Geisinger ja Isberg, 2015). Vaiheen vaihtelu ja siten mahdollisesti CPS-tuotanto on erittäin säänneltyä Ospr-EnvZ-järjestelmässä A., baumannii AB5075, kuten mutaatiot joko OmpR tai EnvZ johti merkittävä kasvu läpinäkymätön tai läpikuultava kytkentä taajuus (Tipton ja Melko, 2017). Vaikka siirtyminen läpikuultava läpinäkymätön johtaa kaksi-kertainen kapselin paksuus, transcriptomic analyyseissä ei ole havaittu mitään eroja ilmaisun tasoa KL geenit kahden vaiheen välillä.
CPS Tavoitteeksi kehittää Rokotteita ja Hoitoja Vastaan A. baumannii
Antibiootti (erityisesti cabapenem) kestävä A., baumannii luokitellaan Maailman terveysjärjestön Priority 1 – kriittiseksi organismiksi uusien mikrobilääkkeiden kehittämiseksi (WHO, 2017). Vaikka ei-antibiootti hoitoja tai rokotteita lisensoitu A. baumannii tällä hetkellä, on lisääntynyt kiinnostus niiden kehittämiseen ja alustavat tutkimukset näyttävät lupaavilta. Pinta-altistuminen ja esiintyvyys patogeenisten kantojen A. baumannii tekee CPS ihanteellinen kohde sekä antimikrobisia hoitoja ja rokotteita (Russo et al., 2013)., Näitä ovat kehitys-vasta-pohjainen hoitojen, kuten ennalta ehkäiseviä rokotteita, passiivinen immunisaatio, ja faagin hoidon (García-Quintanilla et al., 2013).
Useat tutkimukset ovat osoittaneet tehoa passiivinen immunisaatio hiirillä käyttäen CPS-spesifinen vasta-aine, joka on suojaava vastaan bakteeri-haaste 13-55% kliinisistä A. baumannii-isolaattien (Russo et al., 2013; Yang ym., 2017; Lee et al., 2018)., Lisäksi rokotus kanssa konjugaattirokote, joissa CPS glycans kiinnitetty proteiinia harjoittaja saada parempaa immuunijärjestelmän suojaa kuin puhdistettu CPS vastaan monipuolisia A. baumannii-kantoja (Yang et al., 2017).
Kiinnostus faagin hoidon bakteeri-infektioiden hoitoon on lisääntynyt viime vuosina vastauksena kriisiin nousemassa mikrobilääkeresistenssi. Phage-hoito on houkutteleva mahdollinen hoitokanava monilääkeresistenteille A. baumannii-infektioille., Esimerkiksi faagin koodaus CPS depolymerase löytyi hajota CPS noin 10%, neljä 38, kliininen monelle kestävä A. baumannii testattu (Hernandez-Morales et al., 2018). Vaikka isäntä valikoima tämän faagin on rajoitettu, se voi olla sisällytetty osana faagin cocktail maksimoida tehokkuus tai kohdistaa tiettyihin puhkeamisen (Hernandez-Morales et al., 2018). Vaihe, joka valikoivasti katkaisee A: n yhteyden., baumannii CPS klo pseudaminic happo haara voi olla arvokas faagin hoidon, tai tehokkaasti valmistaa puhdistettua CPS rokotteiden ja vasta-aineiden kehittymisen (Lee et al., 2018). Faagin kohdistaminen A. baumannii olivat äskettäin osoitettu olevan stabiili, kun kyllästetty polttaa haavan hoitotuotteet alle erilaisia ehtoja, mukaan lukien läsnäollessa mikrobilääkkeiden (Merabishvili et al., 2017). Kuitenkin, koska suurin osa tutkimuksista tutkii A. baumannii CPS ovat yleensä rajoitu tiettyyn A. baumannii-kantaa, se ei ole tiedossa, jos nämä havainnot kääntää kaikki A., baumannii-isolaatit tai jos ne ovat KanTa-tai kapselityyppisiä. Ymmärtää, mitä rooleja kapseli pelaa useita kantoja on ensiarvoisen tärkeää tunnistaa CPS tyyppejä, jotka edustavat parasta tavoitteet uusien rokotteiden, tai onko mikrobilääkkeiden kehittämistä kohdistaminen kapseli biosynteesin reitti on jopa mahdollista.
Johtopäätökset
Vaikka kapseli on tärkeä virulenssi piirre A. baumannii on olemassa rajoitetusti tietoa saatavilla rooli eri CPS tyypit leikkiä aiheuttaa taudin., Kehittää tehokkaita rokotteita ja hoitoja kohdistaminen CPS, meidän täytyy ensin saada kokonaisvaltaista ymmärrystä mekanismien takana sen synteesi ja ilme, yhdessä etuja että kapseli välittää isäntä bakteerit. Tämä tutkimus on puututtava yhteydessä äärimmäinen vaihtelu CPS serotyyppien löytyy A. baumannii, varmistaa mahdollisten interventioiden työ-kantoja vastaan, jotka tuottavat erilaisia CPS rakenteita., Lisätutkimuksia CPS ovat velvollisia tarjoamaan alustan kehittämiseen ennalta ehkäiseviä toimenpiteitä ja hoitoja vastaan tämä yhä jatkuva ja tappava ihmisen patogeeni.
Kirjailijasuoritukset
JS kirjoitti ensimmäisen luonnoksen. MB antoi akateemisen panoksen ja kriittisen tarkistuksen artikkeliin. FA tuotti genomilähetyksiä ja toimitti kriittisen tarkistuksen käsikirjoitukseen. Kaikki tekijät hyväksyivät lopullisen version.
Rahoitus
Tämän työn tukivat Flinders Medical Research Foundation Myöntää MB. FA: ta tukivat AJ ja IM Naylon ja Playford Trust Ph. D., Apuraha. JS: ää tukivat AJ ja IM Naylon-stipendi.
eturistiriita Lausunto
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
Traub, W. H. (1989). Acinetobacter baumannii serotyypin määritys nosocomial cross-infektion taudinpurkausten selvittämiseksi. J. Clin. Mikrobioli. 27, 2713–2716.
PubMed Abstrakti | Google Scholar
Whitfield, C., ja Paiment, A. (2003)., Ryhmän 1 kapselipolysakkaridien biosynteesi ja kokoaminen Escherichia colissa ja siihen liittyvät solunulkoiset polysakkaridit muissa bakteereissa. Karbohydr. Res. 338, 2491-2502. doi: 10.1016 / j. carres.2003.08.010
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
KUKA (2017). Maailmanlaajuinen prioriteettiluettelo antibiooteille vastustuskykyisistä bakteereista ohjaamaan uusien antibioottien tutkimusta, löytämistä ja kehittämistä. Geneve: Maailman Terveysjärjestö.
Google Scholar