
Bevezetés
a telomerek a lineáris eukarióta kromoszómák végein található nukleoprotein struktúrák. Egyszerre két fő végproblémát oldanak meg. Az első a kromoszóma végvédelméről szól. Becslések szerint a normál emberi sejteknek sejtciklusonként legalább 50 endogén kettős szálú törést (DSBs) kell javítaniuk (Vilenchik and Knudson, 2003)., A telomerek megkülönböztetik a természetes kromoszómális végeket a káros DSB-ktől, és megakadályozzák a méhen kívüli javításukat, például a kromoszómák végponttól végpontig terjedő fúzióival (vanSteensel and deLange, 1997). A második a végreplikációs probléma, amely a megfelelő telomere hossz fenntartásával foglalkozik. Ezt két kutató önállóan ismerte el (Watson, 1972; Olovnikov, 1973). Mivel a replikatív DNS-függő DNS-polimerázok nem képesek a DNS-szintézist a kromoszómák legvégén befejezni, a replikatív telomere-szekvencia veszteség kompenzációjának RNS-függő DNS-polimerázból kell származnia., Ezt az enzimet, az úgynevezett telomerázt, az első telomere minisatellit szekvenciával együtt, a ciliate Tetrahymenában fedezték fel (Blackburn and Gall, 1978; Greider and Blackburn, 1985). Ez azonban csak egy szempont a telomere hossz karbantartás. Terápiás potenciálja (Wellinger et al., 1996; Polotnianka et al., 1998; Pfeiffer and Lingner, 2012).,
a telomeráz, a telomere repeat szekvenciák hozzáadásáért felelős enzim a telomerek 3′ végéhez, egy konzervált komplex enzim, számos összetevővel . Elvileg csak két fő összetevő nélkülözhetetlen a telomeráz enzimaktivitásához, egy katalitikusan aktív fehérje komponens, a telomere reverz transzkriptáz (TERT), valamint egy sablon komponens, amelyet a telomeráz RNS alegység (TR) képez. Míg TERT evolúciósan elég jól konzervált, TR nagyon változó, hosszúságú kezdve ca. 150 nt (Tetrahymena) több mint 2000 nt (gombák nemzetség Neurospora)., Csak egy rövid régió az egész TR molekulában szolgál sablonként az újonnan szintetizált telomere DNS-hez (Greider and Blackburn, 1985; Qi et al., 2013). Ezt a TR régiót általában egy teljes telomere motívum alkotja, amelyet részleges követ, az utóbbi pedig a meglévő telomere DNS lágyító régiójaként szolgál. Bár elvileg csak egyetlen extra nukleotidra van szükség (részleges motívumként), általában egynél több található. Például két extra nukleotid képezi a lágyító motívumot egerekben vagy öt emberben (Blasco et al., 1995; Feng et al., 1995)., A növényekben azonban a sablon régió mérete változó, például kettő az Arabidopsis thaliana-ban, hét az Arabis sp-ben. vagy hat Nicotiana (Fajkus et al., 2019). A többi TR régió strukturális, szabályozási és fehérje interaktív funkciókkal rendelkezik . Lásd még a telomeráz és aktivitási ciklusának vázlatos ábrázolását az 1. ábrán.

1. ábra a telomeráz aktivitási ciklus vázlatos ábrázolása az Arabidopsis típusú telomere sablonnal. TERT, Telomere reverz transzkriptáz; TR, telomeráz RNS alegység., Ábra alapján Sekhri (2014).
milyen változók a Telomere szekvenciák?
a Telomere-szekvenciák általában rövid minisatellitek, jellemzően a képletet követve (TxAyGz)n. a minisatellit-elrendezés abból a módból származik, ahogyan a telomeráz a DNS-t szintetizálja, röviden, többnyire azonos motívumokkal, egyenként., Több hipotézis úgy vélik, hogy egy ilyen megállapodás azért fontos, mert elősegíti a felismerés, a telomer specifikus fehérjék által homo -, illetve heterodimers, valamint a potenciális formában G-quadruplexes, hogy lehet, hogy stabilizálja a kromoszóma vége, vagy szolgálja, mint hordozók telomer-specifikus fehérjék (Spiegel et al., 2020; Tran et al., 2013). A Telomere-szekvenciák jól megőrződnek az evolúció során, a nagy organizmuscsoportok pedig a csoportra jellemző telomere-motívumot használják a telomere DNS felépítéséhez., Fokozatosan növekvő számú vizsgálat és nagy vetítések kimutatták, hogy az összes vizsgált gerinces és sok bazális metazoán TTAGGG-t (Meyne et al., 1989; Traut et al., 2007) míg Euarthropoda (ízeltlábúak), beleértve Hexapoda (rovarok), van TTAGG (Frydrychova et al., 2004; Vitkova et al., 2005). Folyamatosan, számos kivétel halmozódik fel az idő múlásával, például(A (G)1-8) a Dictyosteliumban (Emery and Weiner, 1981), TAGGC in Ascaris lumbricoides (Nematoda) (Muller et al., 1991), Tcagg in Coleoptera (bogarak) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGGG in Giardia (diplomatonads) (Uzlikova et al.,, 2017), vagy Ttnnnnaggg in Yarrowia clade (élesztők) (Cervenak et al., 2019). Ezenkívül a telomeráz-független rendszereket, amelyekben a minisatellite telomere szekvenciát elvesztették és összetett ismétlésekkel helyettesítették, például a Diptera és a Chironomidae (felülvizsgálták (Mason et al., 2016)). Egy általános áttekintést eukarióta telomere szekvencia lásd (Fajkus et al., 2005; Fulneckova et al., 2013).
A Telomere összetétele a növényekben még változatosabb., Itt széles értelemben használjuk a “növények” kifejezést, más néven Archaeplastida vagy kingdom Plantae sensu lato, amely Rhodophyta (vörös algák), Glaucophyta, a Chlorophyte alga fokozat és a Streptophyte alga fokozat (összesen zöld algák néven ismert), valamint Embriophyta (szárazföldi növények) (ezer növényi transzkriptom kezdeményezés, 2019). A tipikus telomere növény szekvencia TTTAGGG, más néven Arabidopsis-típusú (vagy egyszerűen, növényi típusú), mivel felfedezték Arabidopsis thaliana (Richards and Ausubel, 1988), és most sok más faj szinte minden növényrendben., Bár a TTTAGGG még mindig a leggyakoribb, a telomere-szekvenciák jelentős változékonyságot mutatnak a vörös és zöld alga-vonalakban. Ami a vörös algákat (Rhodophyta) illeti, a telomere szekvencia-információk többnyire hiányoznak vagy töredékes, bár néhány telomere jelöltet fedeztek fel silico-ban, például Aatgggggggg a Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT (T) agg for Galdieria sulfuraria (Fulneckova et al., 2013); A TTAGGG a Porphyra umbilicalis (Fulneckova et al., 2013), de további bizonyítékokra van szükség a kromoszómák terminális helyzetének megerősítéséhez., A zöld algák telomere sokfélesége mind a dinamikus változásokat, mind a parafiletikus jellegét tükrözi. Bár a TTTAGGG a Chlorophyta-ban uralkodik, mint például az Ostreococcus nemzetségben (Derelle et al., 2006) és Chlorella (Higashiyama et al., 1995), számos más eltérő motívumot is felfedeztek ott, mint például a TTAGGG a Dunaliella nemzetségben és Stephanosphaeria (Fulneckova et al., 2012), és TTTTAGGG in Chlamydomonas (Petracek et al., 1990). A bazális Streptophyta-ban (Klebsormidiophyceae) a tttaggg-től a TTTTAGGG-ig és a TTTTAGG-ig terjedő motívumok progresszív változásait írták le., A TTAGGG jelenléte Rhodophyta-ban és Glaucophyta-ban arra a hipotézisre vezet, hogy ez az ősi motívum a növényekben (Archaeplastida) (Fulneckova et al., 2013).
a szárazföldi növényeket illetően az egyik első vetítés azt mutatta, hogy az Arabidopsis típusú szekvencia volt a leggyakoribb, és többnyire filogenitásuk révén konzerválódott (Cox et al., 1993; Fuchs et al., 1995), bár ezeknek a szerzőknek egy része már számos kivételt észlelt az Amaryllidaceae családban (korábbi Alliaceae), amelyben az Arabidopsis típusú szekvencia több fajban hiányzott., Később, az első telomere szekvencia szokatlan szárazföldi növények, a gerinces típusú TTAGGG, jellemezte Aloe és néhány más Asparagales (Weiss and Scherthan, 2002; Puizina et al., 2003; Sykorova et al., 2003). A Tttaggg és a TTAGGG telomere szekvencia Aszparagalesben történő ismételt veszteségeiről és megtérüléséről hipotézist fogalmaztak meg (Adams et al., 2001). Az Asparagales rend utófinanszírozásával az APGIII-ban (Angiosperm filogén Csoport 2009) (Bremer et al.,, 2009), kimutatták, hogy a telomere szekvencia-összetételben csak két fő evolúciós kapcsoló fordult elő (több ismétlődő veszteség és nyereség helyett), a következő sorrendben: az első az Iridaceae családban, amelyben a TTTAGGG növénytípusról a ttaggg gerinces típusú TTAGGGRE vált, majd a xeronemataceae, Asphodelaceae és a Core Asparagales (beleértve az Amarillidaceae S.l-t és az Asparagaceae s.l-T.,); és a második az Allioideae alcsaládon belül (korábban külön családként kezelték, Alliaceae), amelyben egy teljesen új telomere-szekvencia alakult ki, CTCGGTTATGG (Fajkus et al., 2016). Az Asparagales-en kívül új telomere-szekvenciákat fedeztek fel olyan szárazföldi növénycsoportokban is, mint az i. Solanaceae, amelyben a Cestrum elegans TTTTTTAGGG telomere-szekvenciáját leírták (Sykorova et al., 2003a; Sykorova et al., 2003b; Peska et al., 2008; Peska et al.,, 2015) és (ii) Lentibulariaceae, ahol genlisea nemzetség mutatott figyelemre méltó sokszínűség néhány faj jellemzi az Arabidopsis típusú telomere ismétlődik, míg mások kiállított keverve szekvencia változatok TTCAGG és TTTCAGG (Tran et al., 2015).
az összes észlelt telomere motívum kivétel ellenére a szárazföldi növények telomer szekvenciáinak valódi sokféleségét valószínűleg nagymértékben alábecsülik. Egy friss kiadvány (Vitales et al.,, 2017), amelyben a szárazföldi növények telomere-szekvenciáinak szűrését végezték, megállapította, hogy a telomere-szekvenciák csak a fajok kevesebb mint 10% – ánál, a növény rDNS-adatbázisában található nemzetségek 40% – ánál ismertek egyértelműen (www.plantrdnadatabase.com), a szárazföldi növényekre vonatkozó molekuláris citogenetikai információkat biztosító forrás (Garcia et al., 2012). A telomere szekvencia eloszlásának összefoglalása növényekben, az APG IV után (az Angiosperm filogén Csoport, 2016) (Byng et al., 2016), valamint a legutóbbi növényi filogén (ezer növényi transzkriptóm kezdeményezés, 2019) A 2.ábrán található.,
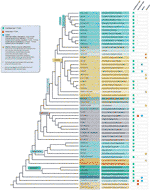
2.ábra telomere motívumok Archaeplastidában (tágabb értelemben vett növények), az APG IV (Az Angiosperm Phylogeny Csoport 2016) és az ezer növényi transzkriptom kezdeményezés (2019) alapján. Az ág hossza nem fejez ki valós idejű skálákat., Az egyszerűség és a helytakarékosság érdekében a fán csillaggal jelölt egyes polifiletikus” csoportokat ” (osztályokat) egyetlen ág képviseli; ugyanezen okból több kisebb sorrendet (amelyek az ábra bal felső részén található Kék négyzetben vannak felsorolva) nem ábrázolnak a fán. Az első tip címke általában növényrendelésekre, néhány esetben osztályzatokra, osztályzatokra, sőt családokra utal; a második címke reprezentatív családokat, néhány esetben reprezentatív megrendeléseket vagy nemzetségeket jelenít meg.
A vetítésektől a felfedezésig: hogyan lehet azonosítani a telomer motívumokat?,
annak bizonyítéka, hogy egy adott jelölt szekvencia valódi telomerikus, több lépést tartalmaz, amelyek megfelelően deklarálják lokalizációját minden kromoszómális terminiben, végül pedig a telomeráz részvételét a szintézisében. Molekuláris citogenetikai (leginkább a Fluoreszcencia in situ Hibridizáció, HAL) vált fontos megjelenítésére, a terminál lokalizáció jelölt szondák jelölt sorozatok egyáltalán kromoszóma található. Azonban önálló hal nem elég bizonyítani a nagyon terminál pozícióját., Például az acepsat356-ot a onionban javasolták telomere jelöltként, a HALELEMZÉS eredményei alapján (Pich and Schubert, 1998). Ennek ellenére az in situ látszólagos terminális helyét soha nem kapcsolták meggyőzően a telomere funkcióhoz. Valójában az Allium minisatellite telomere szekvencia ctcggttatgg és telomerase felfedezése azt jelentené, hogy az AcepSAT356 szubterminális (Fajkus et al., 2019)., A pozitív hal telomer jelek elfedhetik a telomer motívumok apró változásait is, például egy nukleotid polimorfizmusokat, vagy hamis negatív eredmények származhatnak abból, hogy a rövid telomerek a technika kimutatási határa alatt vannak.
két további megközelítés határozza meg a terminális pozíciót nagyobb felbontással, mint a hal; ezek az exonukleáz BAL31 aktivitáson alapulnak., Az első a klasszikus terminális restrikciós fragmentum (TRF) analízis, amelyben a bal31-gyel kezelt minták a terminális fragmensek fokozatos lerövidülését, valamint a jelintenzitás csökkenését mutatják az exonukleáz kezelés idejének növekedésével. A fragmentum hosszának későbbi elemzését Southern-blot hibridizációval végezzük (Fojtova et al., 2015). A második a nem emésztett és a BAL31 emésztett genom DNS összehasonlító genomja (NGS-adatok), párhuzamosan., A bal31 kezelt adatkészletben a telomere szekvenciák jelentős alulreprezentáltak, ezért a terminális szekvenciákat a kezeletlen adatkészlethez képest azonosítjuk, bioinformatikai eszközökkel RepeatExplorer vagy Tandem Repeates Finder.
egy adott telomere szekvencia jelölt másik fontos tesztje egy fajban a telomeráz aktivitásának bizonyítása. Ebben egy hasznos kísérleti megközelítés, amelyet először az emberi sejtek számára fejlesztettek ki, a Telomere Repeat Amplification Protocol (TRAP) (Kim et al., 1994), majd az észlelt termékek szekvenálása (Peska et al.,, 2015; Fajkus et al., 2016), ami egy kicsit kevésbé érzékeny a hamis pozitív eredményekre, mint a hal. Mindezek a módszerek, beleértve a halat (Fuchs et al., 1995; Shibata and Hizume, 2011) és mások, mint a slot-blot hibridizáció (Sykorova et al., 2003c), és TRAP (Fulneckova et al., 2012; Fulneckova et al., 2016), felhasználható a telomerek szűrésére komplex organizmusok széles csoportjain, beleértve a növényeket is., Azonban csak a megfelelően kiválasztott módszerek kombinációja meggyőzően vezethet a jelölt szekvencia telomere funkciójára vonatkozó következtetéshez, mivel az egyetlen megközelítésen alapuló eredmények félrevezetőek lehetnek. A de novo telomere jelölt szekvencia-azonosítási stratégiáinak teljesebb áttekintése, beleértve a Tetrahymena első kísérletét (Greider and Blackburn, 1985), módszertani cikkben foglaljuk össze, hangsúlyt fektetve a rendkívül nagy genomokkal rendelkező növényekben alkalmazott NHS megközelítésre (Peska et al., 2017).
van-e Homoplázia a Telomere-szekvenciákban?,
az ősi telomere szekvenciát TTAGGGNEK tartják, és a leggyakrabban megtalálható az élet fáján (Fulneckova et al., 2013). Mégis, egyértelműnek tűnik, hogy a telomere motívum evolúciójában a homoplázia gyakorisága viszonylag magas. Például a rövid, egyszerű motívumok, mint például a TTTAGGG növénytípus, önállóan és ismételten megjelentek a cryptomonadokban, az oomycete gombákban és az alveolátokban; hasonlóképpen, a gerinces típusú TTAGGG másodlagosan alakult ki bizonyos növénycsoportokban (Asparagales, Rodophyta és Chlorophyta algák) (Sykorova et al., 2003c; Fulneckova et al.,, 2012; Fulneckova et al., 2013; Somanathan és Baysdorfer, 2018). Az oka, hogy a telomer sorozatok jelentek meg gyakrabban, mint a többi, általában bonyolultabb, sorozatok, valószínűleg kapcsolatos szelekciós nyomás, ami szívességet pontosság egy adott szekvencia-specifikus DNS-fehérje kölcsönhatás (Forstemann et al., 2003). Ha minden telomere motívumban változás következik be, a telomer nukleoprotein szerkezetben való interferencia szükségszerűen Genom instabilitáshoz vezet., Ez az oka annak, telomer sorozatok olyan evolúciós stabil, amely nagyon kevés regény, sikeres sorozatok, egy minta megfelel az ötlet, hogy ismételt veszteségek, valamint a megjelenése a jellemző a telomer sorozatok, mint a javasolt Asparagales (Adams et al., 2001).
a telomere-szekvenciákon keresztüli homoplázia megállapítása felveti a kérdést, hogy mik azok a molekuláris okok és folyamatok, amelyek ezekben az eltolódásokban zajlanak? A telomere-szekvencia változása, annak ellenére, hogy bizonyos esetekben triviálisnak tűnik (pl.,, egy extra t), súlyos interferenciát okozhat a genom integritásában, a telomere DNS-fehérje kölcsönhatások zavart egyensúlya miatt. Az sem világos, hogy a változás telomere szekvencia lehet bármilyen evolúciós előnye; ebben a tekintetben, (Tran et al., 2015) azt javasolta, hogy egy “metilálható” citozin megjelenése egy g-gazdag telomere szálban felveti az epigenetikus módosítással történő szabályozás lehetőségét.
milyen molekuláris okai vannak a Telomere motívumok változásainak?,
a telomere szekvencia változásának magyarázatához az első jelölt a telomeráz, a telomeráz RNS (TR) sablon alegysége. A korábban azonosított TR a élesztő gerincesek közé tartozik, hogy egy másik csoport, átiratok, akinek a csatlakozás jellemzője az volt, hogy ők által átírt RNS polimeráz II. (Pol-II.)—minden, de ciliates; ez volt az egyetlen kivétel a Pol-II. átiratok közzététele előtt a föld, növény TR azonosító . Segítségével a viszonylag hosszú telomere motívum Allium keresni a TR belül a teljes RNS szekvencia adatállomány, Fajkus et al., (2019) azt mutatta, hogy az A. thaliana stresszreakciójában korábban jellemzett nem kódoló RNS, az AtR8, valóban a telomeráz RNS alegység (Wu et al., 2012; Fajkus et al., 2019). Az RNS polimeráz III (Pol III) átirata volt, amely a promoter szerkezetében tartalmazza a megfelelő szabályozási elemeket. Hosszú ideig a kutatók azt várták, hogy a növényi TR annyira eltérő lesz, hogy homológiai keresés alapján lehetetlen azonosítani (Cifuentes-Rojas et al., 2011)., Bizonyos fokú hasonlóságot azonban sikeresen alkalmaztak egy közös TR azonosítására több Allium fajban, összehasonlító robbanással. Meglepő, szekvencia homológia, a jelenlét, az azonos szabályozási elemek, valamint a megfelelő sablon régió vezető azonosítása TRs a Allium, Osztályába pedig több mint 70 más távoli rokona növények, beleértve azokat is, akik eltértek a telomer motívumok, mint Genlisea, Cestrum, valamint Tulbaghia., Amennyire tudjuk, még mindig nincs adat semmilyen algal TR-ről, ami tisztázná, hogy a Pol III transzkripciója A TR általános jellemzője minden növény számára, vagy sem. Ez a munka (Fajkus et al., 2019), A CRISPR knock-out és más kísérletek alapján azt is kimutatta, hogy egy korábban azonosított telomeráz RNS-jelölt A. thaliana-ban (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) nem volt a telomeráz funkcionális sablon alegysége, amint azt nem sokkal később (Dew-Budd et al., 2019)., Feltételezve, hogy a Pol II/Pol III függőség a TR transzkripcióhoz megbízható evolúciós marker, a többi fő eukarióta vonal jövőbeli TR kutatása valószínűleg új betekintést nyújt az eukarióták eredetébe. A telomeráz-gének és a telomer-szekvenciák nem ismert információforrások ebben az irányban, és egy Pol III-függő TR biogenezis útvonal megtalálása a ciliate-ben és a növényi vonalokban jelentheti az első lépéseket ebben az irányban (Greider and Blackburn, 1989; Fajkus et al., 2019).
hogyan váltak a kromoszómák Lineárisvá?,
A prokarióták túlnyomó többsége körkörös kromoszómákat tartalmaz, míg az eukarióták szabálya a lineáris kromoszómák., Ezért két lehetséges forgatókönyvet, amely (i) linearizálással által végzett egy primitív telomeráz, előző egyéb folyamatok, ami ahhoz vezetett, hogy a jelenlegi lineáris kromoszóma funkcióit, vagy (ii) linearizálással előre eukarióta körkörös kromoszóma eredetileg telomeráz független, de mielőtt jelenlegi eukaryotes eltértek, egy primitív telomeráz indult, hogy elfoglalja kromoszóma vége lett elengedhetetlen az újonnan alakult lineáris kromoszómák (Nosek et al., 2006). Villasante et al., (2007) olyan evolúciós forgatókönyvet javasolt, amelyben az ősi prokarióta körkörös kromoszóma törése aktivált egy átültetési mechanizmust a DNS végein, lehetővé téve a telomerek rekombinációfüggő replikációs mechanizmus általi kialakulását: ennek a hipotézisnek a következményei meglepő következtetésre vezettek, hogy az eukarióta centromerek telomerekből származnak.
érdekes módon a linearizációval ellentétes folyamat, azaz a körkörös kromoszómák (más néven gyűrűs kromoszómák) kialakulása időről időre felmerült az eukarióták evolúciója során, bár nagyon instabil., Például az Amaranthus tuberculatus esetében a gyűrűs kromoszómák stressz által kiváltott válaszként jelentek meg, amely ellenáll a herbicid (glifozát) ellen; ezek az extra gyűrűs kromoszómák nem mutattak hibridizációt telomere szondákkal a kariotípus-analízisben (Koo et al., 2018). A szinte univerzális telomeráz rendszer és a körkörös kromoszómák kivételessége az eukariótákban nem teszi lehetővé az egyik hipotézis támogatását a másik felett., Azonban a rekombinációs gépek használt alternatív meghosszabbítása telomerek( ALT), egy telomeráz-független út, társított bizonyos emberi rákok (Zhang et al., 2019), már jelen van a prokariótákban. Ezenkívül bizonyíték van arra, hogy a kromoszóma-linearizáció önállóan, különálló prokarióta vonalokban fordul elő (Ferdows and Barbour, 1989; Nosek et al., 1995; Volff és Altenbuchner, 2000). Ezért valószínűbbnek tűnik az a hipotézis, hogy az első lineáris eukarióta kromoszóma (prokarióta ősből származik) telomeráz-független volt., Vannak olyan példák, amelyek azt mutatják, hogy a telomeráz-alapú rendszer nem elengedhetetlen a telomer karbantartás minden eukaryotes: retrotransposons a Drosophila telomerek, műholdas megismétli a Chironomus, egy másik rovar (Rubin, 1978; Biessmann meg Mason, 2003), valamint ALT a telomeráz-negatív emberi rák (Hu et al., 2016; Zhang et al., 2019). Mégis, néhány ilyen rendszerek nem lehet olyan különböző, és talán közös eredete: a Drosophila, a telomere karbantartás, alapuló retrotranszpozíció, nem túl különbözik a telomeráz-alapú mechanizmus (Danilevskaya et al.,, 1998), ami arra a hipotézisre vezet, hogy maga a telomeráz lehet egy korábbi retrotranszpozon. De természetesen a telomeráz-negatív növényfajokat eddig nem fedezték fel, és minden olyan kivételt, amelyben a tipikus növényi típusú telomere hiányzott, később kimutatták, hogy különböző, de még mindig telomeráz-szintetizált motívumok vannak. Mindazonáltal az ALT gép a telomeráz aktivitással párhuzamosan jelen van az üzemekben (Watson and Shippen, 2007; Ruckova et al., 2008). Érdekes kérdések merülnek fel a telomeráz, a telomerek szerepéről és azok növényi daganatokban való fenntartásáról., Vonzó a metasztázis hiánya a növényekben, az ALT jelenléte ellenére, amely talán a növényi szövetek merevségével vagy különböző immunrendszerekkel kapcsolatos, mint az állatoknál (Seyfried and Huysentruyt, 2013).
bár egyre növekvő ismeretekkel rendelkezünk a telomere biológiáról, még mindig nem tudjuk megmagyarázni a telomeráz megjelenését az eukariótákban., A jelenlegi bizonyítékok alátámasztják azt a hipotézist, hogy az eukarióták kialakulása lineáris kromoszómáikkal, teloméráikkal és telomerázukkal együtt összefüggött a spliceoszomális intronok megjelenésével az archaeális gazdaszervezetekben (Koonin, 2006; Fajkus et al., 2019). A TERT és más retroelemek közötti hasonlóságról már egy ideje tárgyalnak (Pardue et al., 1997). Figyelemre méltó, hogy egy viszonylag friss tanulmány kimutatta, hogy a TERT, mint valószínű tagja utódcsoport II intronok, egymás után közel Penelope-szerű elem retrotranszpozonok (Gladyshev and Arkhipova, 2007)., De a TERT csak egyike a két alapvető telomeráz komponensnek, a TR eredete még rejtélyesebb, mivel az összes eukariótában alacsony szekvencia-megőrzés van .
következtetés
a növénygenomika korszakának elején a telomere szekvenciát szinte változatlannak tekintették. A telomerek általános védelme és a telomeráz rendszer azt sugallta, hogy minden növénynek lehet TTTAGGG típusú telomere. A szokatlan telomere-szekvenciák azonosítása komplex növényi genomokban, sok esetben óriási C-értékekkel (például Cestrumban és Allium sp-ben).,), megérte az erőfeszítést, mivel a kivételesen hosszú Allium telomere motívum volt a nyom, amikor valódi TR-t keresett a szárazföldi növényekben. Az újonnan leírt ERP növények, valamint további, a telomer/telomeráz kutatás a bazális clades alga kiderülhet értékes információt arról, hogy a korai fejlődés, ezért a növény telomer kutatás jelentősen hozzájárulnak hipotézisek a megjelenése eukaryotes.
szerzői hozzájárulások
VP és SG egyaránt hozzájárultak e mini-áttekintés írásához, szerkesztéséhez és elkészítéséhez.,
finanszírozás
összeférhetetlenség
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
ezer növényi transzkriptom kezdeményezés (2019). Ezer növényi transzkriptom és a zöld növények filogenomikája. Természet 574 (7780), 679-685. doi: 10.1038 / s41586-019-1693-2
PubMed Abstract / CrossRef teljes szöveg / Google Scholar