
En diploid organisme har to kopier av hvert kromosom. Hvis den har fire kromosomer, det er to par, A og A’ og B og B’, ikke fire ulike kromosomer A, B, C og D. En kopi av hvert kromosom kom fra sin far (for eksempel A og B) og en kopi av hvert kom fra sin mor (f.eks. A’ og B’). Meiose er prosessen med å reduktiv divisjon der en diploid organisme genererer haploid kimceller (i dette tilfellet, med to kromosomer), og hver bakterie celle har én kopi av hvert kromosom., I dette eksemplet, meiose genererer ikke kimceller med En og En’ eller B og B’, snarere er det produserer celler med A og B, eller A og B, eller A’ og B, eller A’ og B’). De homologe kromosomene, som hver består av to søster kromatidene, er koblet sammen i den første fasen av meiose, for eksempel, En med A’ og B med B’ (Figur 8.3; se også Fig. 1.3 og 1.4). Da de homologe kromosomene er flyttet til separate celler på slutten av den første fasen, forsikre at de to homologs ikke bo sammen under reduktiv divisjon i den andre fasen av meiose., Dermed hver bakterie celle mottar haploid komplettere av genetisk materiale, dvs. en kopi av hvert kromosom. Kombinasjonen av to haploid sett av kromosomer under befruktning gjenoppretter diploid state, og syklusen kan fortsette. Unnlatelse av å distribuere en kopi av hvert kromosom til hver bakterie celle har alvorlige konsekvenser. Fraværet av en kopi av et kromosom i en ellers diploid zygote er sannsynlig dødelig. Å ha en ekstra kopi av kromosom (trisomi) fører også til problemer., I mennesker, trisomi for kromosomer 15 eller 18 resultater i perinatal død og trisomi 21 fører til utviklingsmessige feil kjent som «Down’ s syndrom.
Trening 8.2
Hvis dette diploid organisme med kromosomer A, A’, B og B’ gjennomgikk meiose uten homologe sammenkobling og separasjon av homologs til ulike celler, hva brøkdel av den resulterende haploid cellene ville ha en En-type-kromosom (eller’) og et B-type-kromosom (B eller B’)?,
evne til homologe kromosomer å være sammenkoblet under den første fasen av meiose er grunnleggende for å lykkes i denne prosessen, som opprettholder en riktig haploid sett av kromosomer i bakterie celle. Rekombinasjon er en integrert del av sammenkoblingen av homologe kromosomer. Det skjer mellom ikke-søster kromatidene under pachytene fasen av meiose jeg (den første fasen av meiose) og muligens før, når de homologe kromosomene er justert i zygotene (Figur 8.3). Den crossovers av rekombinasjon er synlig i diplotene fase., I løpet av denne fasen, homologe kromosomene delvis separat, men de er fortsatt holdt sammen i ledd heter chiasmata; disse er trolig den faktiske overganger mellom kromatidene av homologe kromosomer. Den chiasmata er gradvis brutt som meiose jeg er fullført, tilsvarende oppløsning av rekombinasjon viderekommende. Under anaphase og telophase av meiose jeg, hver homologe kromosomet flytter til en annen celle, dvs. En og En «i ulike celler, B og B’ i ulike celler i vårt eksempel., Dermed recombinations oppstå i alle meiose, som resulterer i minst en utveksling mellom par homologe kromosomer per meiose.
Nylig genetiske bevis viser at rekombinasjon er nødvendig for homologe sammenkobling av kromosomer under meiose. Genetisk skjermer har avslørt mutanter av gjær og Drosophila som blokkerer sammenkobling av homologe kromosomer. Disse er også defekt i rekombinasjon. Likeledes, mutanter defekt i noen aspekter av rekombinasjon er også defekt i sammenkoblingen., Faktisk, prosessen med synapsis (eller sammenkobling) mellom homologe kromosomer i zygotene, krysset over mellom homologs i pachytene, og oppløsning av crossovere i de siste faser av meiose jeg (diakinesis, metaphase jeg, og anaphase jeg) tilsvarer synapsis, dannelsen av et rekombinant felles og oppløsning som merke progresjon av rekombinasjon, som vil bli forklart nedenfor.
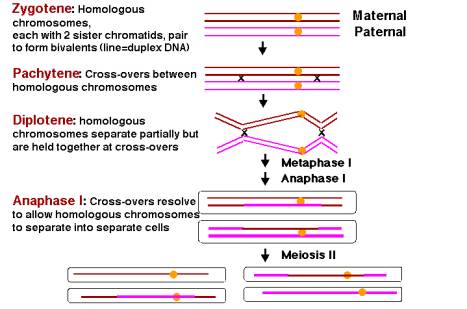
Figur 8.3.Homologe sammenkobling og rekombinasjon i løpet av den første fasen av meiose (meiose jeg)., Etter DNA-syntese er fullført, to eksemplarer av hvert homologe kromosomet er fortsatt koblet på centromeres (gule sirkler). Dette diagrammet starter med identiske kromosomer, referert til som de fire-strand scenen i litteraturen på meiose og rekombinasjon. I denne bruken, hver «strand» er en side av og er en tosidig DNA-molekylet. I dette diagrammet, hver tosidig DNA-molekylet er vist som en enkelt linje, brun for de to søster kromatidene av kromosomet kommer fra moren (mors) og rosa for søster kromatidene fra farsiden kromosom., Bare et homologt par er vist, men det er vanligvis mange flere, f.eks. 4 par kromosomer i Drosophilaand 23 par i mennesker. Under meiose jeg, homologe kromosomene rett inn og deretter separat. På zygotene scenen, de to homologe kromosomene, hver med to søster kromatidene, par langs sin lengde i en prosess som kalles » synapsis. Det resulterer i en gruppe av fire kromatidene kalles en tetrad eller bivalent. Under pachytene, rekombinasjon skjer mellom en mors og en fars side, danner overganger mellom homologe kromosomer., De to homologe kromosomer separat langs mye av sin lengde på diplotene, men de fortsetter å være som holdes sammen ved lokalisert chiasmata, som vises som X-formet strukturer i mikrografer. Disse fysiske koblinger er tenkt å være posisjoner for å krysse over. Under metaphase og anaphase av de første meiotic division, crossovers blir gradvis brutt (med de i endene løst siste) og de to homologe kromosomer (hver fremdeles med to kromatidene ble med på en centromere) er flyttet inn i separate celler., Under den andre meiotic divisjon (meiose II), den centromere av hvert kromosom skiller, slik at de to kromatidene å flytte til egne celler, og dermed endt reduktiv divisjon og gjør fire haploid kimceller.
Bidragsytere og Attribusjoner
-
Ross C. Hardison, T. Ming Chu Professor i Biokjemi og Molekylær Biologi (Pennsylvania State University (universitet)