
Innledning
Telomeres er nucleoprotein strukturer på svært endene av lineær eukaryote kromosomer. De løse to store end-problemer på samme tid. Den første er om kromosom end beskyttelse. Det er anslått at normale menneskelige celler må reparere minst 50 endogene dobbel-strandet pauser (DSBs) per celle per celle-syklus (Vilenchik og Knudson, 2003)., Telomeres skille naturlige kromosomale ender mot skadelige DSBs og hindre deres ektopisk reparasjon, f.eks., av ende-til-ende fusions av kromosomer (vanSteensel og deLange, 1997). Den andre er slutten-replikering problem som omhandler vedlikehold av riktig telomere lengde. Dette ble anerkjent uavhengig av to forskere (Watson, 1972; Olovnikov, 1973). Siden replicative DNA-avhengige DNA polymerases ikke fullstendig DNA-syntese ved svært endene av kromosomene, kompensasjon for replicative telomere sekvens tap må komme fra et RNA-avhengig DNA-polymerase., Dette enzym kalt telomerase, sammen med den første telomere minisatellite sekvens, ble oppdaget i ciliate Tetrahymena (Blackburn og Galle, 1978; Greider og Blackburn, 1985). Imidlertid, dette er bare ett aspekt av telomere lengde vedlikehold. Den epigenetisk regulering av telomere lengde homeostase, inkludert samhandling med lang noncoding telomeric gjenta inneholder RNA og exonuclease aktivitet stier, har også blitt grundig studert på grunn av sin terapeutisk potensial (Wellinger et al., 1996; Polotnianka et al., 1998; Pfeiffer og Lingner, 2012).,
Telomerase, et enzym ansvar for å legge til telomere gjenta sekvenser til 3′ – enden av telomeres, er en bevart enzym komplekse med mange komponenter . I prinsippet er bare to viktigste komponentene er essensielle for telomerase enzymatisk aktivitet, en catalytically aktiv protein komponent, kalt telomere revers transkriptase (TERT), og en mal komponent, dannet av telomerase RNA subunit (TR). Mens TERT er evolusjonært ganske godt bevart, TR er svært variabel, med en lengde som varierer fra ca. 150 nt (Tetrahymena) til mer enn 2000 nt (sopp fra slekten Neurospora)., Bare en kort regionen i hele TR molekyl som fungerer som en mal for nylig syntetisert telomere DNA (Greider og Blackburn, 1985; Qi et al., 2013). Denne regionen i TR er vanligvis dannet av en komplett telomere motiv etterfulgt av en delvis én, sistnevnte tjene som en annealing regionen for eksisterende telomere DNA. Selv om det i prinsippet bare en enkelt ekstra nukleotid er nødvendig (som delvis motiv), vanligvis mer enn én er funnet. For eksempel, to ekstra nukleotider form av avspenning motiv i mus eller fem i menneske (Blasco et al. I 1995; Feng et al., 1995)., I planter, men størrelsen på malen regionen er variabel, for eksempel, to i Arabidopsis thaliana, syv i Arabis sp. eller seks i Nicotiana (Fajkus et al., 2019). Den andre TR regioner har strukturelle, regulatoriske og protein interaktive funksjoner . Se også en skjematisk fremstilling av telomerase og sin aktivitet syklus i Figur 1.

Figur 1 Skjematisk fremstilling av telomerase aktivitet syklus med Arabidopsis-type telomere mal. TERT, Telomere Revers Transkriptase; TR, telomerase RNA subunit., Figur basert på Sekhri (2014).
Hvordan Variabel Er Telomere Sekvenser?
Telomere sekvenser er som regel korte minisatellites tandemly arrangert, vanligvis følgende formel (TxAyGz)n. Den minisatellite ordningen stammer fra måten telomerase syntetiserer DNA, i korte, og for det meste identiske motiver, én etter én., Flere hypoteser anser at en slik ordning er viktig fordi det fremmer anerkjennelse av telomere spesifikke proteiner ved homo – og heterodimers og for potensial til å danne G-quadruplexes som kan stabilisere kromosom ender eller tjene som underlag for telomere-spesifikke proteiner (Spiegel et al., 2020; Tran et al., 2013). Telomere sekvenser er godt bevart gjennom evolusjonen, og store grupper av organismer bruke gruppen-typisk telomere motiv for å bygge sine telomere DNA., Et gradvis økende antall studier og store visninger har vist at alle testet vertebrater og mange basal metazoans bruk TTAGGG (Meyne et al., 1989; Traut et al., 2007), mens Euarthropoda (leddyr), inkludert Hexapoda (insekter), har TTAGG (Frydrychova et al., 2004; Vitkova et al., 2005). Jevnt og trutt, mange unntak akkumuleres over tid, f.eks., (A(G)1-8) i Dictyostelium (Emery og Weiner, 1981), TTAGGC i Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG i Coleoptera (biller) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGGG i Giardia (diplomonads) (Uzlikova et al.,, 2017), eller TTNNNNAGGG i Yarrowia clade (gjær) (Cervenak et al., 2019). Videre, telomerase-uavhengige systemer, som minisatellite telomere sekvensen har blitt borte og erstattet av komplekse gjentar, er representert, for eksempel av Diptera og Chironomidae (anmeldt i (Mason et al., 2016)). For en generell gjennomgang av eukaryote telomere sekvens se (Fajkus et al., 2005; Fulneckova et al., 2013).
Telomere sammensetning i planter er enda mer mangfoldig., Her bruker vi begrepet «planter» i vid forstand, også kjent som Archaeplastida eller rike Plantae sensu lato, og består av Rhodophyta (rød alger), Glaucophyta, den Chlorophyte alger klasse, og det Streptophyte alger klasse (helt kjent som grønn alge), og Embryophyta (landplanter) (Ett Tusen Plante Transcriptomes Initiativ, 2019). Den typiske telomere anlegg sekvensen er TTTAGGG, også kalt Arabidopsis-type (eller rett og slett, plante-type) siden den ble oppdaget i Arabidopsis thaliana (Richards og Ausubel, 1988), og nå i mange andre arter på tvers av nesten alle anlegg bestillinger., Selv om TTTAGGG er fortsatt den mest hyppige, det er betydelig variasjon i telomere sekvenser i rød og grønn alge-linjene. Som for rød alger (Rhodophyta), telomere sekvens informasjon er stort sett mangler eller er fragmentarisk, selv om noen telomere kandidater har blitt oppdaget i silico, for eksempel AATGGGGGG for Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T)AGGG for Galdieria sulphuraria (Fulneckova et al., 2013); TTAGGG har blitt funnet i genomisk leser av Porphyra umbilicalis (Fulneckova et al. I 2013), men flere bevis er nødvendig for å bekrefte sin terminal posisjon på kromosomet., Telomere mangfold i grønne alger gjenspeiler både dynamiske endringer og dens paraphyletic karakter. Selv om TTTAGGG råder i Chlorophyta, for eksempel i slektene Ostreococcus (Derelle et al., 2006) og Chlorella (Nærheten et al., 1995), og mange andre forskjellige motiver har blitt oppdaget der også, slik som TTAGGG i slekten Dunaliella og Stephanosphaeria (Fulneckova et al., 2012), og TTTTAGGG i Chlamydomonas (Petracek et al., 1990). I basal Streptophyta (Klebsormidiophyceae) progressive endringer i motiver fra TTTAGGG å TTTTAGGG og TTTTAGG har blitt beskrevet., Tilstedeværelsen av TTAGGG i Rhodophyta og Glaucophyta fører til hypotesen om at dette er de gamle motiv i planter (Archaeplastida) (Fulneckova et al., 2013).
Om landplanter, en av de første visningene utført viste at Arabidopsis-skriv inn sekvensen var den mest vanlige og var for det meste bevart gjennom sine fylogeni (Cox et al. I 1993; Fuchs et al., 1995), selv om noen av disse forfatterne hadde allerede oppdaget flere unntak i familien Amaryllidaceae (tidligere Alliaceae), der Arabidopsis-skriv inn sekvensen var fraværende i flere arter., Senere, den første telomere sekvens uvanlig for landet planter virveldyr-type TTAGGG, ble preget i Aloe og i enkelte andre Asparagales (Weiss og Scherthan, 2002; Puizina et al., 2003; Sykorova et al., 2003c). En hypotese om gjentatte tap og tilbakeføringer av TTTAGGG og TTAGGG telomere sekvens i Asparagales ble formulert (Adams et al., 2001). Med postrefinement av ordre Asparagales i APGIII (Angiosperm Fylogeni Konsern 2009) (Bremer et al.,, 2009), det viste seg at det bare to store evolusjonære brytere i telomere sekvens sammensetning oppstod (snarere enn flere gjentatte tap og gevinster), i følgende rekkefølge: den første i familien Iridaceae, som et skifte fra plante-type TTTAGGG til virveldyr-type TTAGGG skjedde, etterfulgt av familier Xeronemataceae, Asphodelaceae og kjernen Asparagales (inkludert Amarillidaceae s.l og Asparagaceae s.l.,), og den andre innen subfamily Allioideae (tidligere behandlet som en egen familie, Alliaceae) som en helt ny telomere sekvens dukket opp, CTCGGTTATGGG (Fajkus et al., 2016). Utenfor Asparagales, nye telomere sekvenser har også blitt oppdaget i land anlegg grupper som uensartede som (jeg) Solanaceae, der telomere sekvens av Cestrum elegans TTTTTTAGGG ble beskrevet (Sykorova et al., 2003a; Sykorova et al., 2003b; Peska et al., 2008; Peska et al.,, 2015) og (ii) Lentibulariaceae, hvor slekten Genlisea viste en bemerkelsesverdig mangfold med noen arter preget av Arabidopsis-type telomere gjentar mens andre utstilt blandet rekkefølge varianter TTCAGG og TTTCAGG (Tran et al., 2015).
til Tross for alle telomere motiv unntak oppdaget, det ekte mangfoldet i telomeric sekvenser i landbaserte anlegg er trolig kraftig undervurdert. En nylig publikasjon (Vitales et al.,, 2017), der en screening av land anlegg telomere sekvenser ble utført, fant at telomere sekvenser var bare kjent klart for mindre enn 10% av arter og 40% av arter som finnes i Anlegget rDNA database (www.plantrdnadatabase.com), en ressurs som gir molekylær cytogenetics informasjon om landplanter (Garcia et al., 2012). En oppsummering av telomere sekvens distribusjon i planter, følgende APG IV (Den Angiosperm Fylogeni Gruppe, 2016) (Byng et al., 2016), så vel som det siste anlegget fylogeni (Ett Tusen Plante Transcriptomes Initiativ, 2019) er funnet i Figur 2.,
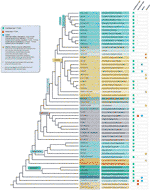
Figur 2 Telomere motiver i Archaeplastida (planter i vid forstand), basert på den APG IV (Den Angiosperm Fylogeni Gruppe 2016) og på den Ene Tusen Plante Transcriptomes Initiative (2019). Gren lengder ikke uttrykk for ekte tidsskalaer., For enkelthets skyld og for å spare plass, visse polyphyletic «grupper» (karakterer) er markert med en stjerne i treet har vært representert ved en enkelt gren; for samme grunn, flere mindre bestillinger (listet opp i den blå firkanten på øvre venstre side av figuren) er ikke avbildet på treet. Den første tips label refererer vanligvis til å plante bestillinger og i noen tilfeller, å divisjoner, karakterer og familier; den andre etiketten viser representant for familier og i et par tilfeller, representant bestillinger eller slekter.
Fra Visningene til Funn: Hvordan Telomeric Motiver Kan Bli Identifisert?,
bevis for at en gitt kandidat sekvensen er en ekte telomeric man inkluderer flere trinn som riktig erklære sin lokalisering på alle kromosomale termini, og til slutt involvering av telomerase i sin syntese. Molekylær cytogenetics (for det meste av Fluorescens in situ Hybridisering, FISK) har blitt viktig for å visualisere terminalen lokalisering av merket prober av kandidat-sekvenser på alle kromosomale termini. Imidlertid, frittstående FISK det er ikke nok til å bevise det veldig terminal posisjon., For eksempel, AcepSAT356 ble foreslått i løk som telomere kandidat, basert på resultater fra FISK analyse (Pich og Schubert, 1998). Likevel, for sin tilsynelatende terminal plassering av in situ har aldri vært overbevisende knyttet til telomere funksjon. Faktisk, oppdagelsen av Allium minisatellite telomere sekvens CTCGGTTATGGG og telomerase ville bety at AcepSAT356 er subterminal (Fajkus et al., 2019)., Positiv FISK telomeric signaler kan også maskere små endringer i telomere motiver som enkelt nukleotid polymorfismer, eller falske negative resultater kan oppstå fra kort telomeres være under deteksjonsgrensen av teknikken.
Det er to flere tilnærminger som bestemmer terminal stilling i større oppløsning enn FISK, og disse er basert på exonuclease BAL31 aktivitet., Den første er den klassiske Terminal Restriction Fragment (TRF) – analyse, der prøvene ble behandlet av BAL31 vis progressive forkortelse av terminal fragmenter og en nedgang i signal intensitet med økende tid av exonuclease behandling. Den påfølgende analyse av fragment lengder er utført av Southern-blot hybridisering (Fojtova et al., 2015). Den andre er komparativ genom skimming (NGS data) av nondigested og BAL31 fordøyd genomisk DNA, i parallell., I BAL31 behandlet dataset, det er en betydelig underrepresentasjon av telomere sekvenser, derfor terminal sekvenser er identifisert ved sammenligning med ubehandlet dataset, ved hjelp av bioinformatikk-verktøy RepeatExplorer eller Tandem Gjentar Finder .
Den andre viktige test av en gitt telomere sekvens kandidat i en art er demonstrasjon av telomerase aktivitet. I denne, en nyttig eksperimentell tilnærming, som ble først utviklet for humane celler, er Telomere Gjenta Forsterkning Protocol (TRAP) (Kim et al., 1994), etterfulgt av sekvensering av de oppdaget produkter (Peska et al.,, 2015; Fajkus et al., 2016), som er litt mindre følsom for falske positive resultater enn FISK. Alle disse metodene, inkludert FISK (Fuchs et al. I 1995; Shibata og Hizume, 2011) og andre som spilleautomat-blot hybridisering (Sykorova et al., 2003c), og FELLE (Fulneckova et al., 2012; Fulneckova et al., 2016), kan brukes til skjermen for telomeres over store grupper av komplekse organismer, inkludert planter., Det er imidlertid bare en kombinasjon av passende valgt metoder kan en overbevisende måte føre til en konklusjon om telomere funksjon av en kandidat sekvens, siden resultatene base på en enkel tilnærming kan være misvisende. En mer fullstendig oversikt over strategier for de novo telomere kandidat sekvens identifikasjon, inkludert den aller første forsøk i Tetrahymena (Greider og Blackburn, 1985) er oppsummert i en metodisk artikkelen, med vekt på NGS tilnærmingen som brukes i anlegg med svært store genomer (Peska et al., 2017).
Er Det Homoplasy i Telomere Sekvenser?,
The ancestral telomere sekvens er tenkt å være TTAGGG og er oftest funnet på tree of life (Fulneckova et al., 2013). Likevel, det synes klart at hyppigheten av homoplasy i telomere motiv utviklingen er relativt høy. For eksempel, korte, enkle motiver som plante-type TTTAGGG har dukket opp uavhengig av hverandre, og flere ganger i cryptomonads, oomycete sopp, og alveolates; på samme måte, virveldyr-type TTAGGG har oppstått sekundært i visse grupper av planter (Asparagales, Rodophyta og Chlorophyta alger) (Sykorova et al., 2003c; Fulneckova et al.,, 2012; Fulneckova et al., 2013; Somanathan og Baysdorfer, 2018). Grunnen til at noen telomere sekvenser har dukket opp oftere enn andre, vanligvis mer komplekse sekvenser er sannsynligvis relatert til utvalget press, noe som ville favør nøyaktighet for en bestemt rekkefølge-spesifikke DNA-protein samhandling (Forstemann et al., 2003). Hvis det var en endring i hvert telomere motiv, forstyrrelser i telomeric nucleoprotein struktur ville nødvendigvis føre til genom instabilitet., Dette er grunnen til telomere sekvenser er så evolusjonær stabil, bestående av svært få ny og vellykket sekvenser, et mønster som er i samsvar med ideen om gjentatte tap og fremveksten av den typiske telomere sekvenser, som er foreslått for Asparagales (Adams et al., 2001).
funn av homoplasy over telomere sekvenser reiser spørsmålet, hva er den molekylære årsaker og prosesser som finner sted i løpet av disse skiftene? En endring i telomere rekkefølge, til tross for tilsynelatende trivielle i noen tilfeller (f.eks., ett ekstra T), kan føre til alvorlige forstyrrelser med genom integritet, på grunn av en forstyrret balansen i telomere DNA-protein interaksjoner. Det er også uklart om en endring i telomere sekvens kan ha noen evolusjonær fordel; i denne forbindelse, (Tran et al., 2015) foreslo at utseendet til en «methylatable» cytosine i en G-rik telomere strand ville øke muligheten for regulering av epigenetisk endring.
Hva Er den Molekylære Årsaker til Endringer i Telomere Motiver?,
for Å forklare telomere sekvensen du vil endre, den første kandidaten er malen subunit av telomerase, telomerase-RNA (TR). Den tidligere identifisert TR fra gjær og vertebrater tilhører en annen gruppe av transkripsjoner, hvis du kobler funksjonen var at de var transkribert av RNA polymerase II (Pol II)—i det hele tatt, men ciliater, og dette pleide å være den eneste unntak fra Pol II transkripsjoner før publisering av landet anlegg TR identifikasjon . Ved å bruke relativt lang telomere motiv av Allium for å lete etter sin TR innen total RNA-sekvens data basseng, Fajkus et al., (2019) viste at en tidligere preget noncoding RNA som er involvert i stress-reaksjon i A. thaliana, kalt AtR8, faktisk var den telomerase RNA subunit (Wu et al., 2012; Fajkus et al., 2019). Det var en transkripsjon av RNA polymerase III (Pol III) inneholder tilsvarende regulatoriske elementer i sin arrangøren struktur. For en lang tid, forskere forventet at anlegget TR ville være så avvikende at det ville være umulig å identifisere den basert på en homologi søk (Cifuentes-Rojas et al., 2011)., Det er imidlertid en viss grad av sammenfall var med hell brukes til å identifisere en felles TR i flere Allium arter med komparative Blast. Overraskende, sekvens homologi, tilstedeværelse av de samme regulatoriske elementer, og en tilsvarende mal for regionen førte til identifisering av TRs i Allium, Arabidopsis og mer enn 70 andre distantly beslektede planter, inkludert de med skilte seg telomere motiver som Genlisea, Cestrum, og Tulbaghia., Så vidt vi vet, det er fortsatt ingen data på alle alge-TR, som ville belyse om Pol III transkripsjon av TR er en generell funksjon for alle planter eller ikke. Dette arbeidet (Fajkus et al., 2019), basert på CRISPR knock-out og andre eksperimenter viste også at en tidligere identifisert telomerase RNA-kandidat i A. thaliana (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) var ikke en funksjonell mal subunit av telomerase, som også ble demonstrert kort tid etter av (Dew-Budd et al., 2019)., Forutsatt at Pol II/Pol III avhengighet for TR transkripsjon er en pålitelig evolusjonære markør, fremtidige TR forskning i andre viktigste eukaryote linjene vil trolig åpne ny innsikt i opprinnelsen av eukaryotes. Telomerase gener og telomere sekvenser er ukjente kilder til informasjon i denne retningen, og funnet av en Pol III avhengige TR biogenese vei i ciliate og anlegg linjene kan representere det første skritt i denne retningen (Greider og Blackburn, 1989; Fajkus et al., 2019).
Hvordan Fikk Kromosomer Bli Lineær?,
Et stort flertall av prokaryotes inneholder sirkulær kromosomer mens lineær kromosomer er regelen i eukaryotes., Derfor er det er to mulige scenarier som enten (i) linearisering ble utført av en primitiv telomerase, før andre prosesser som førte til gjeldende lineært kromosom-funksjoner eller (ii) linearisering av en pre-eukaryote sirkulært kromosom var i utgangspunktet telomerase uavhengige, men like før gjeldende eukaryotes skilte seg, en primitiv telomerase begynte å okkupere kromosom ender, og ble avgjørende for den nyopprettede lineær kromosomer (Nosek et al., 2006). Villasante et al., (2007) har foreslått en evolusjonær scenario der brudd på forfedrenes prokaryotic sirkulært kromosom aktivert en transposition mekanisme på DNA-ender, slik at dannelsen av telomeres av en rekombinasjon-avhengige replikering mekanisme: konsekvensene av denne hypotesen førte til den overraskende konklusjonen at eukaryote centromeres ble hentet fra telomeres.
det er Interessant, det motsatte prosessen for å linearisering, dvs., dannelse av sirkulære kromosomer (også kalt ring kromosomer) har dukket opp fra tid til annen under utviklingen av eukaryotes, selv er svært ustabil., For eksempel, i tilfelle av Amaranthus tuberculatus, ring kromosomer dukket opp som en stress-indusert svar, bærer motstand mot et herbicid (glyfosat); disse ekstra ring kromosomer ikke viser hybridisering med telomere prober i karyotype analyse (Koo et al., 2018). Den nesten universelle telomerase-systemet og exceptionality av sirkulære kromosomer i eukaryotes ikke tillater oss å støtte en hypotese over den andre., Men recombinational maskiner som blir brukt i alternativ forlengelse av telomeres (ALT), en telomerase-uavhengig veien, forbundet med en viss human kreft (Zhang et al., 2019), er allerede til stede i prokaryotes. I tillegg er det bevis på kromosom rettingen skjer uavhengig av hverandre i forskjellige prokaryote linjene (Ferdows og Barbour, 1989; Nosek et al. I 1995; Volff og Altenbuchner, 2000). Derfor hypotesen om at den første lineær eukaryote kromosom (som stammer fra en prokaryote stamfar) var telomerase-uavhengig virker mer sannsynlig., Det er noen eksempler som viser at telomerase-baserte systemet er ikke avgjørende for telomere vedlikehold i alle eukaryotes: retrotransposons i Drosophila telomeres, satellitt gjentas i Chironomus, et annet insekt (Rubin, 1978; Biessmann og Mason, 2003), og ALT i telomerase-negative menneskelige kreft (Hu et al., 2016; Zhang et al., 2019). Likevel, noen av disse systemene kan ikke være så forskjellig, og kan kanskje dele en felles opprinnelse: i Drosophila, telomere vedlikehold, basert i retrotransposition, er ikke så forskjellig fra den telomerase-basert mekanisme (Danilevskaya et al.,, 1998), noe som fører til hypotesen om at telomerase seg selv kan være en tidligere retrotransposon. Men absolutt, telomerase-negative plantearter har ikke blitt oppdaget til dags dato, og alle unntak, som typisk plante-type telomere var fraværende, ble senere vist seg å ha ulike, men likevel telomerase-syntetisert, motiver. Likevel, ALT maskineri er til stede i planter i parallell til telomerase aktivitet (Watson og Shippen, 2007; Ruckova et al., 2008). Interessante spørsmål om hvilken rolle telomerase, telomeres og deres vedlikehold i anlegget svulster oppstår fra den., En attraktiv man er om fravær av metastasering i planter, til tross for tilstedeværelse av ALT, kanskje i slekt med anlegget vev stivhet eller annen immunforsvar enn i dyr (Usa og Huysentruyt, 2013).
Selv om vi er stadig økende kunnskap om telomere biologi, vi er fortsatt i stand til å forklare fremveksten av telomerase i eukaryotes., Nåværende bevis støtter hypotesen om at fremveksten av eukaryotes sammen med sin lineær kromosomer, telomeres, og telomerase var relatert til utseendet av spliceosomal introns i archaeal verter (Koonin, 2006; Fajkus et al., 2019). Likheten mellom TERT og andre retroelements har vært diskutert for noen tid (Pardue et al., 1997). Bemerkelsesverdig er det en relativt fersk studie viste at TERT, som en sannsynlig medlem av avkom gruppe II introns, er sekvensielt nær Penelope-lignende element retrotransposons (Gladyshev og Arkhipova, 2007)., Men TERT er bare en av de to essensielle telomerase komponenter, og TR er i sin opprinnelse, enda mer gåtefull på grunn av sin lave sekvens bevaring tvers av alle eukaryotes .
Konklusjon
I begynnelsen av anlegget genomics era, telomere sekvensen ble ansett som nesten uforanderlig. Den generelle bevaring av telomeres og telomerase system foreslått at alle planter kan ha TTTAGGG plante-type telomere. Identifisering av uvanlig telomere sekvenser i komplekse system genomer, i mange tilfeller med gigantiske C-verdier (for eksempel i Cestrum og Allium sp.,), var verdt innsatsen, siden svært lang Allium telomere motiv var anelse på jakt etter en ekte TR i landet planter. Den nylig beskrevet TR i planter og videre telomere/telomerase forskning innen basal clades av alger kan avsløre verdifull informasjon om tidlig utvikling, derfor plante telomere forskning kan bidra vesentlig til en hypotese om fremveksten av eukaryotes.
Forfatter Bidrag
VP og SG har bidratt like mye til skriving, redigering og klargjøring av denne mini-anmeldelse.,
Midler
interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.
Erkjennelsene
Ett Tusen Plante Transcriptomes Initiative (2019). Tusen plante transcriptomes og phylogenomics av grønne planter. Arten 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar