
introductie
nosocomiale infecties veroorzaakt door multidrugresistente Acinetobacter baumannii worden wereldwijd steeds vaker gebruikt, vooral in de intensive care setting (Wieland et al., 2018). Het succes van deze bacterie wordt vergemakkelijkt door zijn capaciteit om in een verscheidenheid van milieu ‘ s te overleven die door zijn snelle capaciteit wordt verergerd om multidrugweerstand te verwerven. Oppervlakte-koolhydraten spelen een sleutelrol in de algehele conditie en virulentie van A., baumannii (Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Weber et al., 2016). A. baumannii produceert capsulair polysaccharide met een hoog molecuulgewicht (CPS) dat het buitenmembraan omringt (figuur 1) (Russo et al., 2010). Samengesteld uit strak ingepakt het herhalen oligosaccharide subeenheden (eenheden K), vormt CPS een discrete laag op het bacteriële oppervlak die bescherming tegen diverse milieuvoorwaarden verstrekken, die in ontduiking van gastheerimmuunafweer, en stijgende weerstand tegen een aantal antimicrobial samenstellingen (Russo et al., 2010; Iwashkiw et al., 2012; Geisinger and Isberg, 2015).,
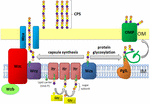
figuur 1. Schematische weergave van capsulepolysaccharide assemblage en export in A. baumannii. De synthese begint met de aanvankelijke transferase (Itr; kastanjebruin) die in het binnenmembraan (IM) wordt gevestigd die de eerste suiker van de zich herhalende eenheid aan een lipidendrager (Und-P; groene cirkel) overbrengen. De volgende suikers worden dan toegevoegd aan de groeiende eenheid door specifieke glycosyltransferases (GTR; geel) aan de cytosolic kant van het binnenmembraan., De subeenheid van de capsule (eenheid K) wordt dan overgebracht naar het periplasma via het geïntegreerde membraaneiwit Wzx (donkerblauw). De suiker-subeenheden worden gepolymeriseerd door het wzy-eiwit (paars) en het WZA/Wzb/WZC-complex (cyaan, kalk, rood) coördineert polymerisatie op hoog niveau en export van de groeiketen, waarbij ze worden verplaatst naar het buitenmembraan (om). Voor glycosylatie koppelt pgll (Oranje) de k-eenheden aan geselecteerde buitenste membraaneiwitten (OMP; donkergroen).
In A. baumannii vindt de capsulemontage en-export plaats via een wzy-afhankelijke route (Hu et al.,, 2013; Kenyon and Hall, 2013; Willis and Whitfield, 2013; Woodward and Naismith, 2016) (figuur 1). Typisch bestaande uit 4-6 suikers, wordt de eenheid K geassembleerd op de lipide dragermolecuul undecaprenylpyrofosfaat (Und-P) dat een steiger voor de groeiende suikerketen verstrekt (Whitfield, 2006). De eerste suiker in de k-eenheid wordt gerekruteerd door een inner membraan (IM)-bound initial transferase (Itr), gevolgd door de sequentiële toevoeging van suikers aan de groeiende K-eenheid door specifieke glycosyltransferase (Gtr) enzymen (figuur 1) (Woodward and Naismith, 2016)., Elke k-eenheid wordt vervolgens overgebracht naar de periplasmische kant van de IM door de WZX-translocase en gepolymeriseerd door Wzy, die de groeiende polysaccharideketen van de ene Und-P-drager naar de volgende inkomende subeenheid (figuur 1) (Collins et al., 2007). Nadat het CPS-polymeer is gesynthetiseerd, wordt het getransporteerd naar het celoppervlak via een sterk gecoördineerd proces waarbij de interactie van drie eiwitten wordt betrokken; Wza, Wzb en Wzc, die de exportmachines omvatten (figuur 1)., De synthese van CPS vertegenwoordigt één arm van een bifurcated weg, aangezien deze eenheden K ook worden gebruikt om bepaalde oppervlakteproteã nen via o-verbonden eiwitglycosylation te versieren (Lees-Miller et al., 2013). In dit geval worden enkele K-eenheden overgedragen aan recipiente eiwitten door de O-oligosaccharyltransferase PglL (figuur 1) (iwashkiw et al., 2012). In A. baumannii draagt eiwitglycosylatie bij aan de vorming van biofilms door de initiële gehechtheid en rijping van biofilms te verbeteren, en pathogeniteit zoals aangetoond in een aantal dierlijke infectiemodellen (iwashkiw et al., 2012; Scott et al., 2014; Harding et al.,, 2015). Biofilm is een de groeistaat waarin de bacteriële gemeenschappen binnen een exopolysaccharidematrijs worden ingesloten en om een significante rol in A. baumannii persistentie en weerstand is getoond te spelen.
andere oppervlakte-koolhydraten waarvan bekend is dat ze de pathogeniteit van A. baumannii beïnvloeden, zijn lipooligosaccharide (LOS) en het exopolysaccharide poly-β-(1-6)-N-acetylglucosamine (PNAG) (Preston et al., 1996; Weber et al., 2016). PNAG vormt de samenhangende “lijm” van biofilms en vormt een aanzienlijk deel van biofilms (Choi et al., 2009; Longo et al., 2014)., In tegenstelling tot de meeste gramnegatieve bacteriën produceert A. baumannii geen traditioneel lipopolysaccharide, maar in plaats daarvan een vergelijkbaar oppervlakte glyco-conjugaat, LOS, bestaande uit een lipide A-kern die geen O-antigeen heeft (Kenyon and Hall, 2013; Kenyon et al., 2014b). Verlies van LOS productie in A. baumannii vermindert de stabiliteit van het buitenmembraan wat leidt tot verminderde fitness (Moffatt et al., 2010; Beceiro et al., 2014).
hoewel veel koolhydraatdelen de pathogeniteit beïnvloeden, kan worden betoogd dat CPS een overheersende virulentiefactor van A. baumannii is. Dit overzicht is bedoeld om te consolideren wat bekend is over A., baumannii capsule met inbegrip van geselecteerde structuren, biosynthese en genorganisatie, de rol van CPS in virulentie, en het potentieel voor CPS als doelwit voor toekomstige vaccin-en geneesmiddelenontwikkeling.
genetische organisatie van K-Loci
naarmate de volledige genomen van meer A. baumannii-isolaten beschikbaar komen, wordt de ware diversiteit van de capsulestructuren binnen deze bacterie duidelijk. Tot op heden zijn meer dan 100 unieke capsule loci (KL) geïdentificeerd in A. baumannii (figuur 2A) (Shashkov et al., 2017). Deze regio ‘ s variëren meestal van 20 tot 35 kb in grootte., Analyse van de genen die CPS-synthese sturen in tien complete genomen van A. baumannii resulteerde oorspronkelijk in de aanwijzing van negen capsuletypes, KL1–KL9, die de basis vormden voor een universeel typschema voor deze loci (Kenyon and Hall, 2013). Deze regeling is vervolgens uitgebreid om de identificatie van nieuwe K-loci mogelijk te maken. De chromosomale locatie van de k-locus, tussen de fkpa-en lldP-genen, wordt sterk behouden tussen A. baumannii-stammen en bevat die genen die nodig zijn voor de biosynthese en export die specifiek zijn voor elk CPS-type (figuur 2) (Hu et al.,, 2013; Kenyon and Hall, 2013). Een uitzondering op deze regel zijn A. baumannii stammen die KL19 en KL39 gebieden bezitten, waar het gen dat codeert voor de wzy polymerase, wzy, wordt gevonden op een klein genetisch eiland elders op het chromosoom (Kenyon et al., 2016a). Bovendien, worden de genen vereist voor sommige van de gemeenschappelijke suikers die in CPS worden gezien elders gevonden., Alle K-loci vertonen een vergelijkbare genetische configuratie, een zeer variabele cluster van synthese-en transferase-genen die nodig zijn voor de biosynthese van unieke KL-type complexe suikers, geflankeerd aan de ene kant door de sterk geconserveerde CPS-exportgenen, en aan de andere kant door een reeks genen die coderen voor geconserveerde enkelvoudige suikers en voorlopers (figuur 2A)., De WZX-en wzy-genen die nodig zijn voor de repeat-unit-verwerking zijn zeer variabel tussen k-loci (figuur 2A, Lichtblauw), wat specificiteit voor bepaalde K-eenheidsstructuren aangeeft en, in het algemeen, de volgorde van gtr-determinanten, die specifieke glycosyltransferasen coderen binnen de KL-regio ‘ s, omgekeerd correspondeert met de volgorde van actie.
het variabele gebied van sommige KL-genclusters, zoals KL37 en KL14, ontberen genen voor complexe suikersynthese, omdat ze alleen eenvoudige suikers in hun K-eenheden bevatten (Arbatsky et al., 2015; Kenyon et al.,, 2015). Ook het toevoegen aan hun diversiteit, bevatten verscheidene KL gebieden overtollige genen die niet voor de synthese van de definitieve K eenheid worden vereist. KL8 en KL9 bevatten bijvoorbeeld twee itr-genen (Kenyon en Hall, 2013) en KL37 heeft pgt1-fosfoglyceroltransferase, maar geen overeenkomstig fosfoglycerolresidu in de bepaalde structuur (Arbatsky et al., 2015). Bovendien onderbreken in KL93 twee elementen van de invoegsequentie (ISAba26 en ISAba22) de pgt1-determinant (Figuur 2) (Kasimova et al., 2017)., De genen die nodig zijn voor specifieke suikerbiosynthese zullen niet worden besproken aangezien deze routes eerder zijn behandeld en buiten het toepassingsgebied van dit overzicht vallen (Hu et al., 2013; Kenyon and Hall, 2013). Bovendien coderen andere genen die zich binnen de KL bevinden producten die worden voorspeld om betrokken te zijn bij acetylering of acylatie modificatie van specifieke glycanen (figuur 2A, roze). Hoewel onderzoek van de k-loci veel kan onthullen over de structuren van de k-eenheid, zijn chemische analyse en biochemische testen nodig om exacte structuren te bepalen en Specifieke verbanden tussen herhaalde suikers te identificeren.,
CPS-structuren
De vroegste studies naar CPS-serotypering van A. baumannii werden gedreven door de noodzaak om een methode te ontwikkelen om isolaten van A. baumannii te onderscheiden van andere Acinetobactersoorten, aangezien fenotypische analyse werd belast met dubbelzinnigheid en verkeerde identificatie (Traub, 1989). Zoals hierboven beschreven, is er fenomenale diversiteit gezien in A. baumannii CPS biosynthese genclusters, die zich vertaalt in de diversiteit gezien in K eenheid structuur (Hu et al., 2013; Kenyon and Hall, 2013). Gezamenlijk meer dan 40 verschillende A., de structuren van de baumannii K eenheid zijn tot nu toe opgehelderd gebruikend NMR spectroscopie en chemische analyse. K eenheidsstructuren verschillen in suikersamenstelling. Zij kunnen derivaten van gemeenschappelijke UDP-gebonden suikers zoals glucose, galactose en glucuronzuur of zeldzame en atypische suikers zoals niet-2-ulosonzuren omvatten. Structuren variëren in lengte en kunnen bestaan uit slechts twee residuen, zoals gezien voor K53 type CPS (Shashkov et al., 2018), of tot vijf of zes monosachariden, zoals die gezien in K37 (figuur 2B) (Arbatsky et al., 2015)., Structuren verschillen ook in de verbindingen zowel binnen als tussen K-eenheden, wat resulteert in de productie van K-eenheden die lineair zijn of zijtakken omvatten, zoals gezien in K1 en K93, respectievelijk (figuur 2B) (Kenyon et al., 2016a; Kasimova et al., 2017). Verschillen in de plaats van specifieke glycosidebindingen en O-acetylatiepatronen van diverse oligosacchariden binnen een structuur dragen ook bij aan de diversiteit van de k-eenheid.,
variatie tussen k-eenheidsstructuren kan subtiel zijn, bijvoorbeeld, K12 en K13 verschillen alleen door de koppeling van twee glycanen, wat het gebruik van een alternatieve wzy-polymerase vereist; dienovereenkomstig zijn de k-loci van beide stammen identiek, behalve voor het wzy-gen (Figuur 2). Als alternatief kan de variatie opvallend zijn, zoals de incorporatie van zeldzame suikers, waaronder pseudaminic, legionaminic, of acinetaminic zuurderivaten zoals waargenomen in respectievelijk K2/6, K49, en K12 / 13 structuren (Figuur 2) (Kenyon et al., 2014a, 2015; Vinogradov et al., 2014; Kasimova et al., 2018)., Interessant is dat acinetaminic zuurderivaten alleen zijn geïdentificeerd in A. baumannii en nergens anders in de natuur worden gevonden (Kenyon et al., 2017). Bovendien bevatten sommige K-eenheden unieke derivaten van specifieke glycanen, bijvoorbeeld het pseudamininezuur van K93 is geacetyleerd met een (R)-3-hydroxybutanoylgroep (Kasimova et al., 2017), terwijl in K2 en K6 het pseudamininezuur niet geacetyleerd is (Figuur 2)., Verder is de structuur van K4 uniek aangezien het slechts aminosugars, d-QuipNAc, en één terminal N-acetyl-D-galactosamine (d-GalpNAcA) vertakking bevat die met een pyruvylgroep wordt afgedekt, een zeldzaam motief en de eerste die in Acinetobacter wordt beschreven (figuur 2A, zwart) (Kenyon et al., 2016b). Naarmate het aantal K-eenheidsstructuren toeneemt, neemt ook het vertrouwen toe om de k-eenheidsstructuur af te leiden uit de analyse van de biosyntheseclusters die in A. baumannii KL zijn geplaatst., Nochtans, hoewel informatief, is een begrip van de rol CPS in pathogenese belangrijk om deze kennis toe te passen om resultaten van A. baumannii besmettingen te verbeteren.
rol bij virulentie, antimicrobiële resistentie en persistentie
Het staat buiten kijf dat de aanwezigheid van CPS essentieel is voor de pathogeniteit van A. baumannii. Niet alleen is het noodzakelijk voor het ontduiken van gastheer immuunafweer (Russo et al., 2010; Geisinger and Isberg, 2015), maar het is belangrijk voor resistentie tegen antimicrobiële verbindingen en overleving in ongunstige omgevingen (Luke et al., 2010; Russo et al.,, 2010; Geisinger en Isberg, 2015). CPS bemiddelt immuunontduiking in veel A. baumannii stammen door de interacties tussen immunogene oppervlaktestructuren van de bacteriën en gastheerafweer te beperken (Preston et al., 1996; Wu et al., 2009; Russo et al., 2010; Umland et al., 2012; Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Wang-Lin et al., 2017). De afschaffing van capsule in meerdere verschillende A. baumannii stammen heeft een verminderde overleving aangetoond in menselijk serum en ascites vloeistof, en verzwakking in ratten en muizen infectie modellen (Russo et al., 2010; Umland et al., 2012; Lees-Miller et al.,, 2013; Sanchez-Larrayoz et al., 2017). Verder verhoogde de up-regulatie van capsuleproductie in de algemeen gebruikte A. baumannii stam ATCC 17978 (K3 CPS type) serumweerstand en virulentie in een muisbesmettings model (Geisinger and Isberg, 2015). Bovendien kunnen nieuwe antimicrobiële behandelingen worden ontwikkeld voor specifieke CPS-typen, bijvoorbeeld die met pseudamininezuur, omdat de aanwezigheid ervan is gecorreleerd met verbeterde virulentie (Hitchen et al., 2010; Kao et al., 2016).
naast bescherming tegen hostafweer, in A., de productie van baumannii CPS verhoogt de weerstand tegen een reeks antimicrobiële verbindingen, waaronder die welke worden gebruikt voor desinfectie in klinische omgevingen (Geisinger and Isberg, 2015; Tipton et al., 2015; Chen et al., 2017). Bovendien beïnvloedt de groei van A. baumannii in subremmende niveaus van antimicrobiële stoffen de productie van CPS. Bijvoorbeeld, leidde de blootstelling aan de antibiotica chloramphenicol of erythromycin tot verbeterde capsulesynthese in ATCC 17978 (Geisinger en Isberg, 2015) en meropenemblootstelling die voor veranderingen wordt geselecteerd die tot een verlies in CPS productie in isolaat 37662 leiden (Chen et al., 2017)., Studies uitgevoerd op een breder scala van A. baumannii stammen zijn nodig om te bepalen of de bescherming tegen antimicrobiële stoffen geboden door CPS is stamspecifiek, capsule-typespecifiek, of universeel.
Het vermogen van A. baumannii om aan te houden in de klinische omgeving heeft ongetwijfeld de kolonisatie en infecties bij gevoelige patiënten versterkt. A. baumannii is in staat om maanden te overleven op ziekenhuisoppervlakken zoals bedrails, meubels en medische hulpmiddelen, waardoor een reservoir dat vaak de bron van transmissie en infectie (Wendt et al., 1997; Gayoso et al., 2013)., De opmerkelijke verdroging tolerantie van A. baumannii wordt verondersteld te zijn te wijten aan een” bust of boom ” strategie, waar een aanhoudende subpopulatie van cellen overleven ten koste van stervende cellen; CPS verhoogt verdroging tolerantie door het verstrekken van een fysieke barrière vergemakkelijken waterretentie (Roberts, 1996; Webster et al ., 2000; Gayoso et al., 2013; Bravo et al., 2016). Een directe rol voor CPS in de droogweerstand werd onlangs aangetoond in de A. baumannii stam AB5075 (K25 CPS type). In deze studie toonde de acapsulaire variant van AB5075 een 2.,5-voudige vermindering van de levensvatbaarheid in vergelijking met de ouderstam (Tipton et al., 2018). Bovendien is bij twee naaste verwanten van A. baumannii, Acinetobacter calcoaceticus en Acinetobacter baylyi, aangetoond dat de productie van exopolysaccharide en/of CPS de overleving van de uitdroging bevordert (Roberson en Firestone, 1992; Ophir en Gutnick, 1994). Naast het beïnvloeden van de weerstand tegen uitdroging, is CPS geassocieerd met andere virulentieeigenschappen, waaronder motiliteit (McQueary et al., 2012; Huang et al., 2014) en de productie van biofilm (Umland et al., 2012; Lees-Miller et al.,, 2013), waardoor zijn rol als pathogene factor wordt versterkt.
recente studies hebben het fasevariabele fenotype van A. baumannii AB5075 in verband gebracht met veranderingen in de CPS-productie, Aangezien zeer virulente ondoorzichtige varianten een CPS-laag produceren met tweemaal de dikte van hun doorschijnende tegenhangers (Chin et al., 2018). Deze overgang van doorschijnend naar ondoorzichtig verhoogde ook dramatisch het pathogene potentieel van A. baumannii AB5075. De resistentie tegen gebruikelijke ziekenhuisdesinfectantia en een subgroep aminoglycoside-antibiotica was ook verhoogd (Tipton et al., 2015; Chin et al.,, 2018) en ondoorzichtige varianten waren ook resistenter tegen humaan lysozym, de cathelicidine-gerelateerde antimicrobiële peptide LL37 en waterstofperoxide in vergelijking met doorschijnende kolonies (Chin et al., 2018). Verder hadden ondoorzichtige isolaten een verhoogde tolerantie voor uitgedroogde omstandigheden en doorschijnende tegenhangers in een muizenbesmettings model (Chin et al., 2018). Aangezien meerdere factoren betrokken zijn bij fasevariatie, werden verdere studies uitgevoerd om de bijdrage van CPS productie aan het meer virulente ondoorzichtige fenotype te bepalen., In een volgende publicatie toonden de auteurs aan dat een acapsulaire variant significant vatbaarder was voor lysozym en desinfecterende middelen in vergelijking met zijn ondoorzichtige ab5075 wild-type ouder (Tipton et al., 2018). Interessant genoeg was er geen verschil in resistentie tegen LL-37 en waterstofperoxide tussen de ondoorzichtige wild-type en acapsulaire stammen, wat suggereert dat andere factoren dan de productie van CPS tot dit fenotype leiden voor ondoorzichtige varianten van A. baumannii (Tipton et al., 2018).,
regulering van de CPS-productie
omgevingsfactoren, zoals temperatuur, osmotische druk en veranderingen in de metaboliet en de beschikbaarheid van ionen, kunnen de CPS-productie van bacteriën beïnvloeden (Hagiwara et al., 2003; Lai et al., 2003; Mouslim et al., 2004; Willenborg et al., 2011). Het is niet verwonderlijk dat er weinig regulerende mechanismen zijn geïdentificeerd voor de productie van CPS, aangezien CPS-niveaus gewoonlijk post-translationeel worden gereguleerd door de fosforylering van CPS-exportmachines (Whitfield and Paiment, 2003; Chiang et al., 2017). In A., baumannii, slechts twee regelgevers van capsuleproductie zijn tot nu toe geïdentificeerd; BfmRS en Ompr-EnvZ, beide twee componenten signaaltransductiesystemen die meerdere regelgevende rollen spelen betrokken bij envelop biogenese (Geisinger and Isberg, 2015; Tipton and Rather, 2017; Geisinger et al., 2018). Wanneer onderworpen aan antibiotische druk, A. baumannii ATCC 17978 CPS expressie werd verhoogd op een bfmrs-afhankelijke manier (Geisinger and Isberg, 2015). Fasevariatie, en dus potentieel CPS-productie, wordt sterk gereguleerd door het OmpR-envz-systeem in A., baumannii AB5075, als mutaties in OmpR of EnvZ resulteerden in een significante toename van de ondoorzichtige tot doorschijnende schakelfrequentie (Tipton and Rather, 2017). Hoewel de overgang van doorschijnend naar ondoorzichtig resulteert in een tweevoudige toename van de capsuledikte, hebben transcriptomic analyses geen verschillen in uitdrukkingsniveaus van kl genen tussen de twee fasen geïdentificeerd.
CPS als doel voor de ontwikkeling van vaccins en Behandelingen tegen A. baumannii
antibioticum (in het bijzonder cabapenem) resistent A., baumannii zijn geclassificeerd als prioriteit 1 van de Wereldgezondheidsorganisatie voor de ontwikkeling van nieuwe antimicrobiële stoffen (WHO, 2017). Hoewel er momenteel geen niet-antibiotische behandelingen of vaccins voor A. baumannii zijn toegelaten, is er een toegenomen interesse in de ontwikkeling ervan en de voorstudies zien er veelbelovend uit. Blootstelling aan het oppervlak en prevalentie in pathogene stammen van A. baumannii maakt CPS een ideaal doelwit voor zowel antimicrobiële behandelingen als vaccins (Russo et al., 2013)., Deze omvatten de ontwikkeling van antilichaam-gebaseerde therapieën zoals profylactische vaccins, passieve immunisatie, en faag therapie (García-Quintanilla et al., 2013).
verschillende studies hebben de werkzaamheid aangetoond van passieve immunisatie bij muizen met een CPS-specifiek antilichaam, dat beschermend is tegen bacteriële challenge met 13-55% van de klinische A. baumannii isolaten (Russo et al., 2013; Yang et al., 2017; Lee et al., 2018)., Bovendien zorgt inoculatie met conjugaatvaccins die CPS glycanen bevatten die aan een eiwitdrager zijn bevestigd voor een betere immuunbescherming dan gezuiverd CPS tegen een breed scala van A. baumannii stammen (Yang et al., 2017).
De interesse in faagtherapie voor de behandeling van bacteriële infecties is de afgelopen jaren toegenomen als reactie op de huidige crisis van toenemende antimicrobiële resistentie. Faag therapie is aantrekkelijk als een potentiële behandeling avenue voor multidrug resistente A. baumannii infecties., Bijvoorbeeld, werd een faag die een CPS depolymerase codeert gevonden om de CPS van ongeveer 10%, vier van 38, klinische multidrug resistente A. baumannii getest (Hernandez-Morales et al., 2018). Hoewel het gastheerbereik van deze faag beperkt is, kan het worden opgenomen als onderdeel van een faagcocktail om de effectiviteit te maximaliseren of specifieke uitbraken te richten (Hernandez-Morales et al., 2018). Een faag die selectief de koppeling van A., baumannii CPS in een pseudaminezuurtak kan waardevol zijn voor faagtherapie, of om efficiënt gezuiverd CPS te produceren voor de ontwikkeling van vaccins en antilichamen (Lee et al., 2018). Faag targeting A. baumannii werden onlangs aangetoond stabiel te zijn wanneer geïmpregneerd in brandwondverzorgingsproducten onder een reeks van voorwaarden, waaronder in de aanwezigheid van antimicrobiële stoffen (Merabishvili et al., 2017). Aangezien de meeste onderzoeken naar CPS van A. baumannii meestal beperkt zijn tot een bepaalde A. baumannii-stam, is het niet bekend of deze bevindingen zich vertalen naar alle A., baumannii isolaten of als ze stam of capsuletype specifiek zijn. Inzicht in welke rollen capsule speelt in meerdere stammen is van het grootste belang bij het identificeren van CPS types die de beste targets voor nieuwe vaccins vertegenwoordigen, of of de ontwikkeling van antimicrobiële gericht capsule biosynthese route is inderdaad zelfs haalbaar.
slotopmerkingen
hoewel capsule een belangrijke virulentieeigenschap van A. baumannii vertegenwoordigt, zijn er beperkte gegevens beschikbaar over de rol die verschillende CPS-typen spelen bij het veroorzaken van ziekte., Om effectieve vaccins en therapieën gericht op CPS te ontwikkelen, moeten we eerst een uitgebreid inzicht krijgen in de mechanismen achter de synthese en expressie ervan, naast de voordelen die capsule aan de gastheerbacteriën overbrengt. Dit onderzoek moet worden aangepakt in de context van de extreme variatie van CPS serotypen gevonden in A. baumannii, om ervoor te zorgen potentiële interventies werken tegen stammen produceren diverse CPS structuren., Verdere studies over CPS zijn nodig om een platform te bieden voor de ontwikkeling van preventieve maatregelen en behandelingen tegen deze steeds hardnekkiger en dodelijke menselijke ziekteverwekker.
Auteursbijdragen
JS schreef het eerste ontwerp. MB leverde academische input en kritische herziening van het artikel. FA produceerde genoomuitlijningen en verstrekte kritische herziening van het manuscript. Alle auteurs hebben de definitieve versie goedgekeurd.
financiering
Dit werk werd ondersteund door een subsidie van de Flinders Medical Research Foundation aan MB. FA werd ondersteund door AJ en IM Naylon en Playford Trust Ph. D., Beurs. JS werd ondersteund door een AJ en IM Naylon beurs.
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Traub, W. H. (1989). Acinetobacter baumannii serotypering voor de afbakening van uitbraken van nosocomiale kruisinfectie. J. Clin. Microbiol. 27, 2713–2716.
PubMed Abstract / Google Scholar
Whitfield, C., and Paiment, A. (2003)., Biosynthese en assemblage van groep 1 capsulaire polysachariden in Escherichia coli en verwante extracellulaire polysachariden in andere bacteriën. Carbohydr. Res.338, 2491-2502. doi: 10.1016 / j.carres.2003.08.010
PubMed Abstract / CrossRef Full Text / Google Scholar
WHO (2017). Global Priority List of antibioticaresistente bacteriën to Guide Research, Discovery, and Development of New antibiotica. Genève: Wereldgezondheidsorganisatie.
Google Scholar