
introduktion
nosokomiala infektioner orsakade av multidrogresistenta Acinetobacter baumannii blir allt vanligare över hela världen, särskilt i intensivvården (Wieland et al., 2018). Framgången för denna bakterie underlättas av dess förmåga att överleva i en mängd olika miljöer som förvärras av dess snabba förmåga att förvärva multidrogresistens. Surface kolhydrater spelar nyckelroller i den övergripande fitness och virulens av A., baumannii (Lees-Miller et al. 2013; för Geisinger och Isberg, 2015; Weber et al., 2016). A. baumannii producerar kapselpolysackarid med hög molekylvikt (CPS) som omger det yttre membranet (Figur 1) (Russo et al., 2010). Består av tätt packade upprepande oligosackarid subenheter (k enheter), CPS bildar ett diskret skikt på den bakteriella ytan som ger skydd mot olika miljöförhållanden, hjälpa till att kringgå värd immun försvar, och ökande resistens mot ett antal antimikrobiella föreningar (Russo et al., 2010; Iwashkiw et al., 2012; Geisinger och Isberg, 2015).,
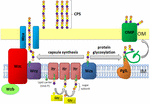
Figur 1. Schematisk representation av kapselpolysackariden montering och export i A. baumannii. Syntes börjar med det initiala transferaset (Itr; maroon) som ligger i det inre membranet (IM) som överför det första sockret i den upprepande enheten till en lipidbärare (Und-p; grön cirkel). Efterföljande sockerarter tillsätts sedan till den växande enheten genom specifika glykosyltransferaser (Gtr; gul) på den cytosoliska sidan av det inre membranet., Kapselsubenheten (k-enheten) överförs sedan till periplasmen via det integrerade membranproteinet Wzx (mörkblå). Sockersubenheterna polymeriseras av Wzy-proteinet (lila) och WZA/Wzb / Wzc-komplexet (cyan, lime, röd) samordnar hög polymerisering och export av den växande kedjan och flyttar dem till det yttre membranet (OM). För glykosylering kopplar PglL (orange) k-enheterna till utvalda yttre membranproteiner (OMP; mörkgrön).
i A. baumannii sker kapselmontering och export via en Wzy-beroende väg (hu et al., 2013; för Kenyon och Hall, 2013; Willis och Whitfield, 2013; Woodward och Naismith, 2016) (Figur 1). Typiskt bestående av 4-6 sockerarter monteras k-enheten på lipidbärarmolekylen undekaprenylpyrofosfat (Und-P) som ger en byggnadsställning för den växande sockerkedjan (Whitfield, 2006). Det första sockret i k-enheten rekryteras av ett inre membran (IM)-bundet initialt transferas (Itr), följt av sekventiell tillsats av sockerarter till den växande k-enheten med specifika glykosyltransferas (Gtr) enzymer (Figur 1) (Woodward och Naismith, 2016)., Varje K enhet överförs sedan till periplasmic sidan av IM den Wzx translokas och polymeriserade genom Wzy, som överför den växande polysackarid kedjan från en Und-P transportören till nästa inkommande subenheten (Figur 1) (Collins et al., 2007). Efter att CPS-polymeren syntetiserats transporteras den till cellytan via en mycket samordnad process som involverar interaktionen mellan tre proteiner; Wza, Wzb och Wzc, som innefattar exportmaskineriet (Figur 1)., CPS-syntes representerar en arm av en bifurcated väg, eftersom dessa k-enheter också används för att dekorera vissa ytproteiner via o-länkad proteinglykosylering (Lees-Miller et al., 2013). I detta fall överförs enstaka k-enheter till mottagarproteiner genom o-oligosackaryltransferas PglL (Figur 1) (Iwashkiw et al., 2012). I A. baumannii bidrar proteinglykosylering till biofilmbildning genom att förbättra initial fastsättning och mognad av biofilmer och patogenicitet som demonstreras i ett antal djurinfektionsmodeller (Iwashkiw et al., 2012; Scott et al., 2014; Harding et al.,, 2015). Biofilm är ett tillväxttillstånd där bakteriella samhällen är inneslutna i en exopolysackaridmatris och har visat sig spela en viktig roll i A. baumannii Persistens och resistens.
andra ytkolhydrater som är kända för att påverka patogeniciteten hos A. baumannii inkluderar lipooligosackarid (LOS) och exopolysackariden poly-β-(1-6)-N-acetylglukosamin (Pnag) (Preston et al., 1996; Weber m.fl., 2016). PNAG bildar ett sammanhängande ” lim ” av biofilmer och utgör en betydande del av biofilmer (Choi et al., 2009; Longo m.fl., 2014)., Till skillnad från de flesta gramnegativa bakterier producerar A. baumannii inte traditionell lipopolysackarid, utan istället en liknande ytglykokonjugat, LOS, bestående av en lipid en kärna som saknar o-antigen (Kenyon och Hall, 2013; Kenyon et al., 2014b). Förlust av LOS-produktion i A. baumannii minskar stabiliteten hos det yttre membranet vilket leder till minskad kondition (Moffatt et al., 2010; Beceiro et al., 2014).
även om många kolhydrater påverkar patogenicitet kan det hävdas att CPS är en dominerande virulensfaktor för A. baumannii. Denna översyn syftar till att konsolidera vad som är känt om A., baumannii kapsel inklusive utvalda strukturer, biosyntes och genorganisation, rollen av CPS i virulens, och potentialen för CPS som ett mål för framtida vaccin och läkemedelsutveckling.
genetisk organisation av K Loci
eftersom de fullständiga genomerna av fler A. baumannii-isolat blir tillgängliga, blir den sanna mångfalden av kapselstrukturer som finns inom denna bakterie uppenbar. Hittills har över 100 unika kapslar loci (KL) identifierats i A. baumannii (figur 2a) (Shashkov et al., 2017). Dessa regioner varierar vanligtvis från 20 till 35 kb i storlek., Analys av generna som styr CPS-syntes i tio kompletta A. baumannii-genomer resulterade ursprungligen i beteckningen av nio kapseltyper, KL1-KL9, som blev grunden för ett universellt typsystem för dessa loci (Kenyon och Hall, 2013). Detta system har därefter utvidgats för att möjliggöra identifiering av nya k loci. Kromosomavvikelser plats av K-locus, mellan fkpA och lldP gener, är starkt bevarad mellan A. baumannii stammar och innehåller de gener som krävs för biosyntes och export som är specifika för varje CPS-typ (Figur 2) (Hu et al., 2013; för Kenyon och Hall, 2013). Ett undantag till denna regel är A. baumannii stammar som har KL19 och KL39 regioner, där den gen som kodar för den Wzy-polymeras, wzy, finns på en liten genetisk ön på andra ställen på kromosom (Kenyon et al., 2016a). Dessutom finns de gener som krävs för några av de vanliga sockerarterna som ses i CPS någon annanstans., Alla k loci visar en liknande genetisk konfiguration, en mycket varierande kluster av syntes och transferasgener som krävs för biosyntesen av unika KL-typ komplexa sockerarter, flankerad på ena sidan av de högt konserverade CPS export gener, och på andra sidan av en uppsättning gener som kodar konserverade enkla sockerarter och prekursorer (figur 2A)., De wzx-och wzy-gener som krävs för upprepad bearbetning är mycket varierande mellan k loci (figur 2a, ljusblå), vilket indikerar specificitet för särskilda k-enhetsstrukturer och i allmänhet ordningen för gtr-determinanter, som kodar för specifika glykosyltransferaser inom KL-regionerna, motsvarar omvänt handlingsordningen.
variabelområdet för vissa KL-genkluster, såsom KL37 och KL14, saknar gener för komplex sockersyntes, eftersom de bara innehåller enkla sockerarter i sina k-enheter (Arbatsky et al. År 2015; Kenyon et al.,, 2015). Också lägga till deras mångfald, flera KL regioner innehåller överflödiga gener som inte krävs för syntesen av den slutliga k-enheten. Till exempel KL8 och KL9 innehåller två itr gener (Kenyon och Hall, 2013) och KL37 har pgt1 phosphoglyceroltransferase, men ingen motsvarande phosphoglycerol rester i bestämd struktur (Arbatsky et al., 2015). Dessutom, i KL93, två styck sekvens element (ISAba26 och ISAba22) avbryta pgt1 faktor (Figur 2) (Kasimova et al., 2017)., De gener som krävs för specifik sockerbiosyntes kommer inte att diskuteras eftersom dessa vägar har täckts tidigare och ligger utanför ramen för denna översyn (hu et al. 2013; för Kenyon och Hall, 2013). Dessutom kodar andra gener inom KL-produkterna för produkter som förutspås vara involverade i acetylering eller acyleringsmodifiering av specifika glykaner (figur 2a, rosa). Även om undersökning av k-loci kan avslöja mycket om k-enhetsstrukturer krävs kemisk analys och biokemisk testning för att bestämma exakta strukturer och identifiera specifika kopplingar mellan upprepade sockerarter.,
CPS-strukturer
de tidigaste studierna på CPS-serotypning av A. baumannii drevs av behovet av att utveckla en metod för att diskriminera A. baumannii-isolat från andra Acinetobacter-arter, eftersom fenotypisk analys belastades med tvetydighet och misidentifiering (Traub, 1989). Som beskrivits ovan, Det finns fenomenal mångfald ses i A. baumannii CPS biosyntes genkluster, som översätter till den mångfald som ses i k enhetsstruktur (hu et al. 2013; för Kenyon och Hall, 2013). Tillsammans över 40 olika A., baumannii k-enhetsstrukturer har hittills belysts med hjälp av NMR-spektroskopi och kemisk analys. K enhetsstrukturer skiljer sig åt i sockerkompositionen. De kan innefatta derivat av vanliga UDP-länkade sockerarter såsom glukos, galaktos och glukuronsyra eller sällsynta och atypiska sockerarter såsom icke-2-ulosonsyror. Strukturerna varierar i längd och kan bestå av endast två restprodukter, som sett för K53 typ CPS (Shashkov et al., 2018), eller upp till fem eller sex monosackarider, såsom de som ses i K37 (Figur 2B) (Arbatsky et al., 2015)., Strukturer skiljer sig också i kopplingarna både inom och mellan k-enheter vilket resulterar i produktion av k-enheter som är linjära eller involverar sidogrenar, vilket ses i K1 respektive K93 (Figur 2B) (Kenyon et al., 2016a; Kasimova et al., 2017). Skillnader i placeringen av specifika glykosidbindningar och O-acetyleringsmönster av olika oligosackarider inom en struktur bidrar också till k-enhetens mångfald.,
variationen mellan k-enhetsstrukturer kan vara subtil, till exempel skiljer sig K12 och K13 endast genom kopplingen mellan två glykaner, vilket kräver användning av ett alternativt Wzy-polymeras; följaktligen är K-loci av båda stammarna identiska med undantag för wzy-genen (Figur 2). Alternativt kan variationen vara slående, såsom inkorporering av sällsynta sockerarter inklusive pseudaminsyra, legionaminsyra eller acinetaminsyraderivat enligt K2/6, K49 respektive K12/13 strukturer (Figur 2) (Kenyon et al., 2014a, 2015; Vinogradov et al., Och med 2014. Kasimova et al., 2018)., Intressant nog har acinetaminsyraderivat endast identifierats i A. baumannii och finns ingen annanstans i naturen (Kenyon et al., 2017). Dessutom innehåller vissa k-enheter unika derivat av specifika glykaner, till exempel är pseudaminsyra av K93 acetylerad med en (R)-3-hydroxybutanoyl-grupp (Kasimova et al., 2017), medan i k2 och K6 är pseudaminsyran icke-acetylerad (Figur 2)., Dessutom, struktur K4 är unikt eftersom det innehåller bara aminosugars, D-QuipNAc, och en terminal N-acetyl-D-galaktosamin (D-GalpNAcA) filial som är förseglat med ett pyruvyl grupp, en sällsynt motiv och den första som kommer att beskrivas i Acinetobacter (Figur 2A, svart) (Kenyon et al., 2016b). Som antalet k-enhetsstrukturer klarlagda ökar, så gör förtroendet att dra slutsatsen k-enhetsstrukturen från analys av biosynteskluster placerade i A. baumannii KL., Men även informativ, en förståelse för den roll CPS spelar i patogenesen är viktigt att tillämpa denna kunskap för att förbättra resultaten av A. baumannii infektioner.
roll i virulens, antimikrobiell resistens och persistens
det är utan tvekan att förekomsten av CPS är avgörande för A. baumannii patogenicitet. Inte bara är det nödvändigt för undvikande av värd immunförsvar (Russo et al., 2010; Geisinger och Isberg, 2015), men det är viktigt för resistens mot antimikrobiella föreningar och överlevnad i ogynnsamma miljöer (Luke et al., 2010, Russo m.fl.,, 2010; Geisinger och Isberg, 2015). CPS medierar immun skatteflykt i många A. baumannii stammar genom att begränsa interaktioner mellan immunogena ytstrukturer av bakterier och värd försvar (Preston et al., 1996; Wu m.fl., 2009; Russo m.fl., 2010; Umland et al., 2012; Lees-Miller et al. 2013; för Geisinger och Isberg, 2015, Wang-Lin et al., 2017). Avskaffandet av kapsel i flera olika A. baumannii stammar har visat minskad överlevnad i humant serum och ascites vätska, och dämpning i råtta och murina infektion modeller (Russo et al., 2010; Umland et al., 2012; Lees-Miller et al., 2013; för Sanchez-Larrayoz et al., 2017). Dessutom ökade uppregleringen av kapselproduktionen i den vanligen använda A. baumannii-stammen ATCC 17978 (K3 CPS-typen) serumresistens och virulens i en musinfektionsmodell (Geisinger och isberg, 2015). Dessutom kan nya antimikrobiella behandlingar utvecklas för specifika CPS-typer, till exempel de som innehåller pseudaminsyra, eftersom dess närvaro har korrelerats med ökad virulens (Hitchen et al., 2010, Kao m.fl., 2016).
förutom skydd mot värdförsvar, i A., baumannii CPS-produktionen ökar resistensen mot en rad antimikrobiella föreningar, inklusive de som används för desinfektion i kliniska miljöer (Geisinger och isberg, 2015; Tipton et al. År 2015; Chen et al., 2017). Vidare påverkar tillväxten av A. baumannii i subhämmande nivåer av antimikrobiella medel CPS-produktionen. Till exempel exponering för antibiotika kloramfenikol eller erytromycin lett till ökad kapsel syntes i ATCC 17978 (Geisinger och Isberg, 2015) och meropenem exponering utvalda för mutationer som leder till en förlust i CPS produktion i isolera 37662 (Chen et al., 2017)., Studier utförda på ett bredare spektrum av A. baumannii-stammar krävs för att identifiera om skyddet mot antimikrobiella medel som CPS erbjuder är stamspecifikt, kapseltyp specifikt eller universellt.
förmågan hos A. baumannii att kvarstå i den kliniska miljön har utan tvekan ökat kolonisering och infektioner hos mottagliga patienter. A. baumannii kan överleva i månader på sjukhusytor som sängskenor, möbler och medicintekniska produkter, vilket ger en reservoar som ofta är källan till överföring och infektion (Wendt et al. 1997; Gayoso m.fl., 2013)., Den anmärkningsvärda uttorkningstoleransen hos A. baumannii tros bero på en” byst eller boom ” – strategi, där en beständig subpopulation av celler överlever på bekostnad av döende celler; CPS förbättrar torkningstolerans genom att tillhandahålla en fysisk barriär som underlättar vattenretention (Roberts, 1996; Webster et al., 2000; Gayoso m.fl., 2013; Bravo et al., 2016). En direkt roll för CPS i torkningsbeständighet visades nyligen i A. baumannii-stammen AB5075 (K25 CPS-typen). I denna studie visade den akapsulära varianten av AB5075 en 2.,5-faldig minskning av lönsamheten jämfört med föräldrastammen (Tipton et al., 2018). Dessutom, i två nära släktingar till A. baumannii, Acinetobacter calcoaceticus och Acinetobacter baylyi, produktion av exopolysaccharide och/eller CPS har visat att främja uttorkning överlevnad (Roberson och Firestone, 1992; Ophir och Gutnick, 1994). Förutom att påverka resistens mot uttorkning har CPS associerats med andra virulensdrag, inklusive motilitet (McQueary et al., 2012; Huang et al., 2014) och produktion av biofilm (Umland et al., 2012; Lees-Miller et al.,, 2013), vilket cementerar sin roll som en patogen faktor.
nyligen genomförda studier har kopplat den fasvariabla fenotypen av A. baumannii AB5075 med förändringar i CPS-produktion, eftersom mycket virulenta ogenomskinliga varianter producerar ett CPS-lager med dubbelt tjockleken hos sina genomskinliga motsvarigheter (Chin et al., 2018). Denna övergång från genomskinlig till ogenomskinlig ökade också dramatiskt den patogena potentialen hos A. baumannii AB5075. Resistens mot vanliga sjukhus desinfektionsmedel och en delmängd av aminoglykosidantibiotika ökade också (Tipton et al. År 2015, Haka et al.,, 2018) och ogenomskinliga varianter var också mer resistenta mot humant lysozym, den katelicidinrelaterade antimikrobiella peptiden LL37 och väteperoxid jämfört med genomskinliga kolonier (Chin et al., 2018). Dessutom hade ogenomskinliga isolat en ökad tolerans mot uttorkade tillstånd och out-tävlade genomskinliga motsvarigheter i en mus infektion modell (Chin et al., 2018). Eftersom flera faktorer är involverade i fasvariation utfördes ytterligare studier för att bestämma CPS-produktionens bidrag till den mer virulenta ogenomskinliga fenotypen., I en följande publikation visade författarna att en akapsulär variant var signifikant mer mottaglig för lysozym och desinfektionsmedel jämfört med dess ogenomskinliga ab5075 wild-type förälder (Tipton et al., 2018). Intressant var det ingen skillnad i resistens mot LL-37 och väteperoxid mellan de ogenomskinliga vildtypen och acapsulära stammarna, vilket tyder på andra faktorer än CPS-produktion leder till denna fenotyp för ogenomskinliga varianter av A. baumannii (Tipton et al., 2018).,
reglering av CPS-produktion
miljömässiga signaler, såsom temperatur, osmotiskt tryck och förändringar i metabolit och jontillgänglighet kan påverka bakteriell CPS-produktion (Hagiwara et al., 2003; lai m.fl., 2003; Mouslim et al., 2004; Willenborg m.fl., 2011). Det är inte förvånande att få regleringsmekanismer har identifierats för CPS-produktion, eftersom CPS-nivåerna vanligtvis regleras efter translationellt genom fosforylering av CPS-exportmaskiner (Whitfield och Paiment, 2003; Chiang et al., 2017). I A., baumannii, bara två regulatorer av kapsel produktionen har hittills identifierats; BfmRS och OmpR-EnvZ, både två-komponent signal transduktion system som spelar flera olika lagstadgade roller som är involverade i kuvert biogenes (Geisinger och Isberg, 2015; Tipton och Snarare 2017; Geisinger et al., 2018). Vid exponering för antibiotikatryck ökade A. baumannii ATCC 17978 CPS-uttryck på ett bfmrs-beroende sätt (Geisinger och isberg, 2015). Fasvariation, och därmed potentiellt CPS-produktion, är starkt reglerad av OmpR-envz-systemet i A., baumannii AB5075, som mutationer i antingen OmpR eller EnvZ resulterade i en betydande ökning i ogenomskinliga till genomskinligt switching frekvens (Tipton och Snarare 2017). Även om övergången från genomskinlig till ogenomskinlig resulterar i en tvåfaldig ökning av kapseltjockleken, har transkriptomiska analyser inte identifierat några skillnader i uttrycksnivåer av KL-gener mellan de två faserna.
CPS som mål för utveckling av vacciner och behandlingar mot A. baumannii
antibiotikum (speciellt cabapenem) resistent A., baumannii klassificeras en Världshälsoorganisation Priority 1 kritisk organism för utveckling av nya antimikrobiella medel (WHO, 2017). Även om det inte finns några icke-antibiotika behandlingar eller vacciner som är licensierade för A. baumannii för närvarande, finns det ett ökat intresse för deras utveckling och preliminära studier ser lovande ut. Ytexponering och prevalens i patogena stammar av A. baumannii gör CPS till ett idealiskt mål för både antimikrobiella behandlingar och vacciner (Russo et al., 2013)., Dessa inkluderar utveckling av antikroppsbaserade terapier som profylaktiska vacciner, passiv immunisering och fagbehandling (García-Quintanilla et al., 2013).
Flera studier har visat att effekten av passiv immunisering i möss med hjälp av en CPS-specifik antikropp, som är skyddande mot bakteriell utmaning med 13-55% av kliniska A. baumannii isolat (Russo et al., 2013; Yang et al., 2017; Lee et al., 2018)., Dessutom framkallar inokulering med konjugatvacciner som innehåller CPS-glykaner kopplade till en proteinbärare bättre immunskydd än renat CPS mot ett varierat utbud av A. baumannii-stammar (Yang et al., 2017).
intresset för fagterapi för att behandla bakterieinfektioner har ökat de senaste åren som svar på den nuvarande krisen med stigande antimikrobiell resistens. Phage therapy är attraktivt som en potentiell behandlingsväg för multidrugresistenta A. baumannii-infektioner., Till exempel befanns en fag som kodade en CPS-depolymeras försämra CPS av cirka 10%, fyra av 38, klinisk multidrugresistent A. baumannii testad (Hernandez-Morales et al., 2018). Även om värdområdet för denna FAG är begränsat kan det införlivas som en del av en fagcocktail för att maximera effektiviteten eller rikta specifika utbrott (Hernandez-Morales et al., 2018). En fag som selektivt klyver kopplingen av A., baumannii CPS vid en pseudaminsyragren kan vara värdefull för fagterapi, eller för att effektivt tillverka renade CPS för vaccin-och antikroppsutveckling (Lee et al., 2018). Phage targeting A. baumannii visade sig nyligen vara stabil när den impregnerades i brännsårvårdsprodukter under en rad förhållanden, inklusive i närvaro av antimikrobiella medel (Merabishvili et al., 2017). Men eftersom majoriteten av studier som undersöker A. baumannii CPS vanligtvis är begränsade till en viss A. baumannii-stam, är det inte känt om dessa fynd översätter till alla A., baumannii isolat eller om de är stam eller kapsel typ specifik. Att förstå vilka roller kapsel spelar i flera stammar är avgörande för att identifiera CPS-typer som representerar de bästa målen för nya vacciner, eller om utvecklingen av antimikrobiella medel som riktar sig till kapselbiosyntesvägen verkligen är genomförbar.
avslutande anmärkningar
även om kapseln representerar ett viktigt virulensdrag hos A. baumannii finns det begränsade data tillgängliga om vilken roll olika CPS-typer spelar för att orsaka sjukdom., För att utveckla effektiva vacciner och terapier som riktar sig till CPS måste vi först få en omfattande förståelse för mekanismerna bakom syntes och uttryck, tillsammans med de fördelar som kapsel förmedlar till värdbakterierna. Denna forskning måste behandlas i samband med den extrema variationen av CPS-serotyper som finns i A. baumannii, för att säkerställa potentiella insatser mot stammar som producerar olika CPS-strukturer., Ytterligare studier om CPS krävs för att tillhandahålla en plattform för utveckling av förebyggande åtgärder och behandlingar mot denna alltmer ihållande och dödliga mänskliga patogen.
Författarbidrag
JS skrev det första utkastet. MB gav akademisk input och kritisk översyn av artikeln. FA producerade genomjusteringar och gav kritisk översyn av manuskriptet. Alla författare godkände den slutliga versionen.
finansiering
detta arbete stöddes av ett Flinders Medical Research Foundation bidrag till MB. FA stöddes av AJ och IM Naylon och Playford Förtroende Ph. D., Stipendier. JS var stöds av en AJ och IM Naylon Stipendium.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Traub, W. H. (1989). Acinetobacter baumannii serotypning för avgränsning av utbrott av nosokomial korsinfektion. J. Clin. Microbiol. 27, 2713–2716.
PubMed Abstract/Google Scholar
Whitfield, C. Och Paiment, A. (2003)., Biosyntes och montering av kapselpolysackarider i Grupp 1 i Escherichia coli och relaterade extracellulära polysackarider i andra bakterier. Karbohydr. Res. 338, 2491-2502. doi: 10.1016 / j. carres.2003.08.010
PubMed Abstrakt | CrossRef Full Text | Google Scholar
SOM (2017). Global prioriteringslista över antibiotikaresistenta bakterier för att styra forskning, upptäckt och utveckling av nya antibiotika. Geneva: Världshälsoorganisationen.
Google Scholar