
introduktion
telomerer är nukleoproteinstrukturer i slutet av linjära eukaryota kromosomer. De löser två stora slutproblem samtidigt. Den första handlar om kromosomslutskydd. Det uppskattas att normala mänskliga celler måste reparera minst 50 endogena dubbelsträngade raster (Dsb) per cell per cellcykel (Vilenchik och Knudson, 2003)., Telomerer skiljer de naturliga kromosomändarna från skadliga Dsb och förhindrar deras ektopisk reparation, t. ex. genom end-to-end-fusioner av kromosomer (vanSteensel och deLange, 1997). Den andra är slut-replikering problem som behandlar upprätthållandet av rätt telomere längder. Detta erkändes självständigt av två forskare (Watson, 1972; Olovnikov, 1973). Eftersom replikativa DNA-beroende DNA-polymeraser inte kan komplettera DNA-syntesen i kromosomernas ändar, måste ersättning för replikativ telomeresekvensförlust komma från ett RNA-beroende DNA-polymeras., Detta enzym, kallat telomeras, tillsammans med den första telomere minisatellitsekvensen, upptäcktes i ciliate Tetrahymena (Blackburn och Gall, 1978; Greider och Blackburn, 1985). Detta är dock bara en aspekt av telomere längd underhåll. Den epigenetiska regleringen av telomerelängdshemostas, inklusive interaktion mellan långa icke-kodande telomerisk upprepning innehållande RNA och exonukleasaktivitetsvägar, har också studerats i stor utsträckning på grund av dess terapeutiska potential (Wellinger et al., 1996; Polotnianka m.fl., 1998; Pfeiffer och Lingner, 2012).,
telomeras, det enzym som ansvarar för att lägga till telomeres repetersekvenser till 3-änden av telomerer, är ett konserverat komplext enzym med många komponenter . I princip är endast två huvudkomponenter väsentliga för telomeras enzymatisk aktivitet, en katalytiskt aktiv proteinkomponent, kallad telomere omvänd transkriptas (TERT) och en mallkomponent, bildad av telomeras RNA-subenheten (TR). Medan TERT är evolutionärt ganska väl bevarad, är TR mycket variabel, med längder från ca. 150 nt (Tetrahymena) till mer än 2,000 nt (svampar från släktet Neurospora)., Endast en kort region i hela TR-molekylen tjänar som en mall för nysyntetiserat telomere DNA (Greider och Blackburn, 1985; Qi et al., 2013). Denna region i TR bildas vanligtvis av ett fullständigt telomere-motiv följt av en partiell, den senare tjänar som en glödgning region för det befintliga telomere-DNA. Även om det i princip bara behövs en enda extra nukleotid (som ett partiellt motiv), finns vanligtvis mer än en. Till exempel bildar två extra nukleotider det glödande motivet hos möss eller fem hos människa (Blasco et al., 1995; Feng et al., 1995)., I växter är dock mallregionens storlek variabel, t. ex. två i Arabidopsis thaliana, sju i Arabis sp. eller sex i Nicotiana (Fajkus et al., 2019). De andra TR-regionerna har strukturella, reglerande och proteininteraktiva funktioner . Se även en schematisk bild av telomeras och dess aktivitetscykel i Figur 1.

Figur 1 Schematisk representation av telomeras aktivitetscykel med Arabidopsis-typ telomere Mall. Tert, Telomere omvänt transkriptas; TR, telomeras rna subenhet., Figur baserad på Sekhri (2014).
hur variabel är Telomeresekvenser?
Telomere sekvenser är vanligtvis korta minisatelliter tandemly arrangerade, typiskt efter formeln (TxAyGz)n. minisatellit arrangemanget härstammar från det sätt på vilket telomeras syntetiserar DNA, kort sagt, och mestadels identiska motiv, en efter en., Flera hypoteser anser att ett sådant arrangemang är viktigt eftersom det främjar erkännandet av telomerespecifika proteiner av homo – och heterodimerer och för potentialen att bilda G-quadruplexes som kan stabilisera kromosomändar eller fungera som substrat för telomerespecifika proteiner (Spiegel et al., 2020; Tran m.fl., 2013). Telomeresekvenser är väl bevarade genom evolution, och stora grupper av organismer använder gruppen-typiska telomere motiv att bygga sin telomere DNA., Ett gradvis ökande antal studier och stora screenings har visat att alla testade ryggradsdjur och många basala metazoaner använder TTAGGG (Meyne et al. 1989; Traut m.fl., 2007) medan Euarthropoda (leddjur), inklusive Hexapoda (insekter), har TTAGG (Frydrychova et al., 2004; Vitkova et al., 2005). Stadigt, många undantag är ackumuleras över tid, t ex, A(G)1-8) i Dictyostelium (Emery och Weiner, 1981), TTAGGC i Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG i Coleoptera (skalbaggar) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGGG i Giardia (diplomonads) (Uzlikova et al.,, 2017), eller TTNNNNAGGG i Yarrowia klad (jäst) (Cervenak et al., 2019). Dessutom representeras telomerasoberoende system, där minisatellit telomere-sekvensen har förlorats och ersatts av komplexa upprepningar, till exempel av Diptera och Chironomidae (granskad i (Mason et al., 2016)). För en allmän översyn av eukaryotisk telomeresekvens se (Fajkus et al., 2005; Fulneckova m.fl., 2013).
Telomere sammansättning i växter är ännu mer varierande., Här använder vi termen ”växter” i bred bemärkelse, även känd som Archaeplastida eller rike Växtriket bemärkelse lato, och som består av Rhodophyta (röda alger), Glaucophyta, den Chlorophyte alger klass och Streptophyte alger klass (tillsammans kallas för ” gröna alger), och Embryophyta (mark, växter) (Tusen Växt Transcriptomes Initiativ, 2019). Den typiska telomere växtföljden är TTTAGGG, även kallad Arabidopsis-typ (eller helt enkelt, växt-typ) eftersom den upptäcktes i Arabidopsis thaliana (Richards och Ausubel, 1988) och nu i många andra arter över nästan alla växtorder., Även om TTTAGGG fortfarande är den vanligaste, finns det betydande variationer i telomeresekvenser i röda och gröna alglinjer. När det gäller rödalger (Rhodophyta) saknas eller fragmenteras telomeresequentin oftast, även om vissa telomerekandidater har upptäckts i silico, såsom Aatgggggggg för Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T)AGGG för Galdieria sulphuraria (Fulneckova et al., 2013); TTAGGG har hittats i genomisk läser av Porphyra umbilicalis (Fulneckova et al., 2013), men mer bevis behövs för att bekräfta deras terminala position på kromosomer., Telomeres mångfald i grönalger återspeglar både dynamiska förändringar och dess parafyletiska karaktär. Även om TTTAGGG råder i Chlorophyta, såsom i släkten Ostreococcus (Derelle et al., 2006) och Chlorella (Higashiyama et al., 1995), många andra olika motiv har upptäckts där också, sådana som TTAGGG i släktet Dunaliella och Stephanosphaeria (Fulneckova et al., 2012), och TTTTAGGG i Chlamydomonas (Petracek et al., 1990). I basal Streptophyta (Klebsormidiophyceae) progressiva förändringar i form av motiv från TTTAGGG att TTTTAGGG och TTTTAGG har beskrivits., Förekomsten av TTAGGG i Rhodophyta och Glaucophyta leder till hypotesen att detta är det historiska motiv i växter (Archaeplastida) (Fulneckova et al., 2013).
När det gäller markplantor visade en av de första screeningarna att Arabidopsis-typsekvensen var den vanligaste och var mestadels bevarad genom deras fylogeni (Cox et al., 1993; Fuchs et al., 1995), även om vissa av dessa författare redan hade upptäckt flera undantag i familjen Amaryllidaceae (tidigare Alliaceae), där Arabidopsis-typsekvensen var frånvarande i flera arter., Senare karakteriserades den första telomeresekvensen ovanlig för markväxter, ryggradsdjur-typen TTAGGG, i Aloe och i några andra Asparagales (Weiss och Scherthan, 2002; Puizina et al., 2003; Sykorova et al., 2003c). En hypotes om upprepade förluster och återvinningar av TTTAGGG och TTAGGG dna-sekvens i Asparagales formulerades (Adams et al., 2001). Med postrefinement av ordningen Asparagales i APGIII (Gömfröiga växter Fylogeni Koncernen 2009) (Bremer et al.,, 2009), visade det sig att endast två stora evolutionära växlar i dna-sekvensen sammansättning skett (i stället för flera upprepade förluster och vinster), i följande ordning: den första i familjen Iridaceae, där en övergång från växt-typ TTTAGGG till ryggradsdjur-typ TTAGGG hänt, följt av familjer Xeronemataceae, Asphodelaceae och kärnan Asparagales (inklusive Amarillidaceae s.l och Asparagaceae s.l.,); och den andra inom underfamiljen Allioideae (tidigare behandlad som en separat familj, Alliaceae) där en helt ny telomeresekvens uppstod, CTCGGTTATGGG (Fajkus et al., 2016). Utanför Asparagales har nya telomeresekvenser också upptäckts i markväxtgrupper som olikartade som i) Solanaceae, där telomeresekvensen av Cestrum elegans TTTTTTAGGG beskrevs (Sykorova et al., 2003a; Sykorova et al., 2003b; Peska m.fl., 2008; Peska m.fl.,, År 2015) och (ii) Lentibulariaceae, där släktet Genlisea visade en anmärkningsvärd mångfald med vissa arter kännetecknas av Arabidopsis-typ telomer upprepar medan andra uppvisade samsas sekvens varianter TTCAGG och TTTCAGG (Tran et al., 2015).
trots alla de telomere motiv undantag upptäcks, den verkliga mångfalden i telomera sekvenser i markplantor är förmodligen kraftigt underskattas. En ny publikation (Vitales et al.,, 2017), där en screening av mark växt telomere sekvenser utfördes, fann att telomere sekvenser endast var kända tydligt för mindre än 10% av arten och 40% av släktena som finns i växt rDNA-databasen (www.plantrdnadatabase.com), en resurs som tillhandahåller molekylär cytogenetik information om markväxter (Garcia et al., 2012). En sammanfattning av telomere sequence distribution i växter, efter APG IV (The Angiosperm Phylogeny Group, 2016) (Byng et al., 2016), liksom den senaste anläggningen fylogeny (tusen Plant Transcriptomes Initiative, 2019) finns i Figur 2.,
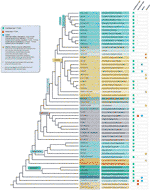
Figur 2 Telomere motiv i Archaeplastida (växter i vid bemärkelse), baserat på APG IV (Angiosperm Phylogeny Group 2016) och på One Thousand Plant Transcriptomes Initiative (2019). Grenlängder uttrycker inte realtidsskalor., För enkelhet och för att spara utrymme har vissa polyfyletiska ”grupper” (betyg) markerade med en asterisk i trädet representerats av en enda gren; av samma anledning är flera mindre order (listade i den blå kvadraten på vänster övre sida av figuren) inte avbildade på trädet. Den första tipsetiketten hänvisar vanligtvis till växtorder och i några fall till divisioner, betyg och till och med familjer.den andra etiketten visar representativa familjer och i några fall representativa order eller släkten.
från Visningar till upptäckt: hur Telomera motiv kan identifieras?,
beviset på att en given kandidatsekvens är en riktig telomerisk innehåller flera steg som korrekt förklarar lokaliseringen vid alla kromosomala termini och så småningom involvering av telomeras i syntesen. Molekylär cytogenetik (mestadels genom fluorescens i situ hybridisering, fisk) har blivit viktigt för att visualisera terminal lokalisering av märkta sonder av kandidatsekvenser på alla kromosomala termini. Fristående fisk är det dock inte tillräckligt att bevisa själva terminalpositionen., Till exempel föreslogs AcepSAT356 i onion som telomere-kandidat, baserat på resultat från FISH analysis (Pich och Schubert, 1998). Dess skenbara terminalplats vid in situ har dock aldrig varit övertygande kopplad till telomere-funktionen. Faktiskt, upptäckten av Allium minisatellite dna-sekvens CTCGGTTATGGG och telomeras skulle innebära att AcepSAT356 är subterminal (Fajkus et al., 2019)., Positiva fisk telomera signaler kan också maskera små förändringar i telomere motiv som enda nukleotid polymorfismer, eller falskt negativa resultat kan bero på korta telomerer är under detektionsgränsen för tekniken.
det finns ytterligare två tillvägagångssätt som bestämmer terminalpositionen vid större upplösning än fisk; dessa är baserade på exonukleasbal31-aktivitet., Den första är den klassiska Terminalrestriktionsfragmentet (TRF) – analysen, där prover behandlade med BAL31 visar progressiv förkortning av terminalfragment och en minskning av signalintensiteten med ökande tid för exonukleasbehandling. Den efterföljande analysen av fragmentlängder utförs av Southern-blot hybridisering (Fojtova et al., 2015). Den andra är jämförande genomet skimming (NGS data) nondigested och BAL31 rötas genomiskt DNA, parallellt., I BAL31-behandlade dataset finns en signifikant underrepresentation av telomeresekvenser, därför identifieras terminalsekvenserna i jämförelse med den obehandlade datauppsättningen med hjälp av bioinformatics tools RepeatExplorer eller Tandem Repeats Finder .
det andra viktiga testet av en viss telomeresekvenskandidat i en art är demonstrationen av telomerasaktivitet. I detta är ett användbart experimentellt tillvägagångssätt, utvecklat först för mänskliga celler, Telomere Repeat Amplification Protocol (TRAP) (Kim et al., 1994), följt av sekvensering av de upptäckta produkterna (Peska et al., År 2015; Fajkus et al., 2016), vilket är lite mindre känsligt för falskt positiva resultat än fisk. Alla dessa metoder, inklusive FISK (Fuchs et al., 1995; Shibata och Hizume, 2011) och andra som slot-blot hybridisering (Sykorova et al., 2003c), och FÄLLA (Fulneckova et al., 2012; Fulneckova et al., 2016), kan användas för att Skärm för telomerer över breda grupper av komplexa organismer, inklusive växter., Endast en kombination av lämpligt valda metoder kan dock på ett övertygande sätt leda till en slutsats om en kandidatsekvens telomerefunktion, eftersom resultatbasen på ett enda tillvägagångssätt kan vara vilseledande. En mer fullständig översikt över strategierna för de novo telomere kandidatsekvens identifiering, inklusive det allra första försöket i Tetrahymena (Greider och Blackburn, 1985) sammanfattas i en metodologisk artikel, med betoning på NGS-metoden som används i växter med extremt stora genom (Peska et al., 2017).
finns det homoplasi i Telomeresekvenser?,
ancestral telomere-sekvensen tros vara TTAGGG och är den vanligast förekommande över livets träd (Fulneckova et al., 2013). Ändå verkar det tydligt att frekvensen av homoplasi i telomeres motivutveckling är relativt hög. Till exempel, korta, enkla motiv som växt-typ TTTAGGG har dykt upp självständigt och upprepade gånger i cryptomonads, oomycete svampar, och kryptomonader, likaså ryggradsdjur-typ TTAGGG har framkommit i andra hand i vissa grupper av växter (Asparagales, Rodophyta och Chlorophyta alger) (Sykorova et al., 2003c; Fulneckova m.fl.,, 2012; Fulneckova et al. 2013; för Somanathan och Baysdorfer, 2018). Anledningen till att vissa telomeresekvenser har uppstått oftare än andra, är vanligtvis mer komplexa sekvenser troligen relaterade till urvalstryck, vilket skulle gynna noggrannhet för en viss sekvensspecifik DNA-proteininteraktion (Forstemann et al., 2003). Om det fanns en förändring i varje telomere-motiv, skulle interferens i telomerisk nukleoproteinstruktur nödvändigtvis leda till genominstabilitet., Detta är anledningen till att telomere sekvenser är så evolutionära stabila, innefattande mycket få nya och framgångsrika sekvenser, ett mönster som överensstämmer med tanken på upprepade förluster och uppkomsten av de typiska telomeresekvenserna, som föreslagits för Asparagales (Adams et al., 2001).
upptäckten av homoplasi över telomeresekvenser väcker frågan, vilka molekylära orsaker och processer som sker under dessa skift? En förändring i telomere-sekvensen, trots skenbara triviala i vissa fall (t. ex.,, en extra T), kan orsaka allvarliga störningar med genomintegritet, på grund av en störd balans i telomerens DNA-proteininteraktioner. Det är också oklart om en förändring i telomere-sekvensen kan ha någon evolutionär fördel ;i detta avseende, (Tran et al., 2015) föreslog att utseendet av en ”metylatable” cytosin i en g-rik telomere-sträng skulle öka möjligheten till reglering genom epigenetisk modifiering.
vilka är de molekylära orsakerna till förändringar i Telomeremotiven?,
för att förklara telomere-sekvensändring är den första kandidaten mallunderenheten för telomeras, telomeras RNA (TR). Den tidigare identifierade TR från jäst och ryggradsdjur hör till en annan grupp av transkript, vars anslutande funktion var att de transkriberades av RNA polymeras II (Pol II) – i alla utom ciliater; detta brukade vara det enda undantaget från Pol II-transkript före publicering av markplant TR-identifieringen . Genom att använda alliums relativt långa telomeremotiv för att leta efter sitt TR inom den totala rna-sekvensdatapoolen, Fajkus et al., (2019) visade att ett tidigare karakteriserat icke-kodande RNA som var involverat i stressreaktionen i A. thaliana, kallat AtR8, verkligen var telomeras RNA-subenheten (Wu et al., 2012; Fajkus et al., 2019). Det var ett transkript av RNA-polymeras III (Pol III) som innehöll motsvarande regulatoriska element i dess promotorstruktur. Under lång tid förväntade forskarna att plant TR skulle vara så divergent att det skulle vara omöjligt att identifiera det baserat på en homologisökning (Cifuentes-Rojas et al., 2011)., En viss grad av likhet användes emellertid framgångsrikt för att identifiera ett gemensamt TR i flera Alliumarter med jämförande Blast. Överraskande, sekvenshomologi, närvaron av samma regulatoriska element och en motsvarande mallregion ledde till identifiering av TRs i Allium, Arabidopsis och mer än 70 andra distansrelaterade växter, inklusive de med divergerade telomere motiv som Genlisea, Cestrum och Tulbaghia., Såvitt vi vet finns det fortfarande inga data om någon algal TR, vilket skulle belysa huruvida Pol III transkription av TR är en allmän egenskap för alla växter eller inte. Detta arbete (Fajkus et al., 2019), baserat på CRISPR knock-out och andra experiment visade också att ett tidigare identifierat telomeras RNA-kandidat i A. thaliana (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) var inte en funktionell Mall subenhet av telomeras, vilket också visades kort efter av (Dew-Budd et al., 2019)., Om man antar att Pol II/Pol III-beroendet för TR-transkription är en tillförlitlig evolutionär markör, kommer framtida TR-forskning i andra viktiga eukaryotiska linjer förmodligen att öppna nya insikter om ursprunget för eukaryoter. Telomeras-gener och telomere-sekvenser är okända informationskällor i denna riktning, och upptäckten av en Pol III-beroende TR-biogenesväg i ciliat och växtlinjer kan representera de första stegen i denna riktning (Greider och Blackburn, 1989; Fajkus et al., 2019).
Hur blev kromosomerna linjära?,
en stor majoritet av prokaryoter innehåller cirkulära kromosomer medan linjära kromosomer är regeln i eukaryoter., Därför finns det två möjliga scenarier där antingen (i) linearisering utfördes av ett primitivt telomeras, före andra processer som ledde till nuvarande linjära kromosomala egenskaper och funktioner eller (ii) linearisering av en pre-eukaryotisk cirkulär kromosom var ursprungligen telomeras oberoende, men strax före nuvarande eukaryoter divergerade, en primitiv telomeras började ockupera kromosom ändar och blev avgörande för de nybildade linjära kromosomerna (Nosek et al., 2006). Villasante et al., (2007) föreslog ett evolutionärt scenario där brytningen av den förfäders prokaryotiska cirkulära kromosomen aktiverade en införlivandemekanism vid DNA-ändar, vilket möjliggör bildandet av telomerer genom en rekombinationsberoende replikeringsmekanism: konsekvenserna av denna hypotes ledde till den överraskande slutsatsen att eukaryotiska centromerer härleddes från telomerer.
intressant är att den motsatta processen till linearisering, dvs bildandet av cirkulära kromosomer (även kallade ringkromosomer) har uppstått från tid till annan under utvecklingen av eukaryoter, även om de är mycket instabila., Till exempel, när det gäller Amaranthus tuberculatus, uppträdde ringkromosomer som ett stressinducerat svar, som bär motstånd mot en herbicid( glyfosat); dessa extra ringkromosomer visade inte hybridisering med telomereprober i karyotypanalysen (Koo et al., 2018). Det nästan universella telomerassystemet och undantagsordningen för cirkulära kromosomer i eukaryoter tillåter oss inte att stödja en hypotes över den andra., Den rekombinationella maskiner som används i den alternativa förlängningen av telomerer (ALT), en telomerasoberoende väg, associerad med vissa mänskliga cancerformer (Zhang et al., 2019), är redan närvarande i prokaryotes. Dessutom finns det bevis på kromosomlinjärisering som uppträder oberoende i distinkta prokaryotlinjer (Ferdows och Barbour, 1989; Nosek et al., 1995; Volff och Altenbuchner, 2000). Därför verkar hypotesen att den första linjära eukaryotiska kromosomen (som härrör från en prokaryote förfader) var telomerasoberoende mer sannolikt., Det finns några exempel som visar att det telomerasbaserade systemet inte är nödvändigt för telomere-underhåll i alla eukaryoter: retrotransposons i Drosophila telomeres, satellitrepetitioner i Chironomus, en annan insekt (Rubin, 1978; Biessmann och Mason, 2003) och ALT i telomeras-negativa mänskliga cancerformer (hu et al., 2016; Zhang et al., 2019). Ändå kan vissa av dessa system inte vara så olika, och kan kanske dela ett gemensamt ursprung: i Drosophila är telomere-underhållet, baserat i retrotransposition, inte alltför distinkt från den telomerasbaserade mekanismen (Danilevskaya et al.,, 1998), vilket leder till hypotesen att telomeras själv kan vara en tidigare retrotransposon. Men visst har telomeras-negativa växtarter hittills inte upptäckts och alla undantag, där den typiska växttypen telomere var frånvarande, senare visade sig ha olika men fortfarande telomeras-syntetiserade motiv. Ändå är ALT-maskinen närvarande i växter parallellt med telomerasaktiviteten (Watson och Shippen, 2007; Ruckova et al., 2008). Intressanta frågor om rollen som telomeras, telomerer och deras underhåll i växttumörer uppstår från det., En attraktiv handlar om frånvaron av metastaser i växter, trots närvaron av ALT, kanske relaterad till växtvävnadsstyvhet eller olika immunsystem än hos djur (Seyfried och Huysentruyt, 2013).
även om vi får ökad kunskap om telomerebiologi, kan vi fortfarande inte förklara uppkomsten av telomeras i eukaryoter., Aktuella bevis stöder hypotesen att uppkomsten av eukaryoter tillsammans med sina linjära kromosomer, telomerer och telomeras var relaterad till uppkomsten av spliceosomal introner i archaeal värdar (Koonin, 2006; Fajkus et al., 2019). Likheten mellan TERT och andra retroelements har diskuterats en tid (Pardue et al., 1997). Det är anmärkningsvärt att en relativt nyligen genomförd studie visade att TERT, som en trolig medlem i avelsgrupp II introner, jämfört med föregående kvartal är nära till Penelope-liknande element retrotransposons (Gladyshev och Arkhipova, 2007)., Men TERT är bara en av de två väsentliga telomeraskomponenterna, och TR är, i sitt ursprung, ännu mer gåtfull på grund av dess låga sekvensbevarande över alla eukaryoter .
slutsats
i början av växtgenomiktiden ansågs telomeresekvensen nästan förändrad. Det allmänna bevarandet av telomerer och telomeras-systemet föreslog att alla växter kan ha tttaggg-anläggningstypen telomere. Identifiering av ovanliga telomeresekvenser i komplexa växtgenom, i många fall med jätte C-värden (såsom i Cestrum och Allium sp.,), var värt ansträngningen, eftersom det exceptionellt långa Allium telomere motivet var ledtråden i att leta efter en äkta TR i markplantor. Den nyligen beskrivna TR i växter och ytterligare telomere / telomeras forskning i basala klader av alger kan avslöja värdefull information om tidig utveckling, därför växt telomere forskning kan avsevärt bidra till hypoteser om uppkomsten av eukaryoter.
författarens bidrag
VP och SG har bidragit lika till skrivning, redigering och förberedelse av denna mini-översyn.,
finansiering
intressekonflikt
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
ett tusen växt Transcriptomes Initiative (2019). Tusen växt transkriptomer och fylogenomik av gröna växter. Natur 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstrakt | CrossRef Full Text | Google Scholar