
Indledning
Nosokomielle infektioner forårsaget af multidrug resistente Acinetobacter baumannii bliver mere og mere udbredt på verdensplan, især i intensiv indstilling (Wieland et al., 2018). Succesen med denne bakterie lettes af dens evne til at overleve i en række forskellige miljøer forstærket af dens hurtige evne til at erhverve multidrugsresistens. Overflade kulhydrater spiller en central rolle i den generelle kondition og virulens af A., baumannii (Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Geeber et al., 2016). A. baumannii producerer kapselpolysaccharid med høj molekylvægt (CPS), der omgiver den ydre membran (Figur 1) (Russo et al., 2010). CPS består af tætpakkede gentagne oligosaccharidunderenheder (k-enheder) og danner et diskret lag på bakterieoverfladen, der giver beskyttelse mod forskellige miljøforhold, hjælper med unddragelse af værtens immunforsvar og øger modstanden mod en række antimikrobielle forbindelser (Russo et al., 2010; i .ashki.et al., 2012; Geisinger og Isberg, 2015).,
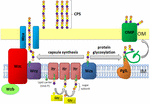
Figur 1. Skematisk repræsentation af kapselpolysaccharid samling og eksport i A. baumannii. Syntese begynder med den indledende transferase (Itr; maroon) placeret i den indre membran (IM), der overfører det første sukker af den gentagne enhed til en lipidbærer (Und-p; grøn cirkel). Efterfølgende sukker tilsættes derefter til den voksende enhed ved specifikke glycosyltransferaser (GTR; gul) på den cytosoliske side af den indre membran., Kapselunderenheden (k-enhed) overføres derefter til periplasm via det integrerede membranprotein… (Mørkeblå). Sukkerunderenhederne polymeriseres af protein proteiny-proteinet (lilla), og complex .a /b .b/. .c-komplekset (cyan, lime, red) koordinerer polymerisation på højt niveau og eksport af den voksende kæde og flytter dem til den ydre membran (OM). Til glykosylering forbinder PglL (orange) k-enhederne med udvalgte ydre membranproteiner (OMP; mørkegrøn).
I A. baumannii, kapsel forsamling og eksport sker via en Wzy-afhængig vej (Hu et al.,, 2013; Kenyon og Hall, 2013; Willis og Whitfield, 2013; Woodward og Naismith, 2016) (Figur 1). Typisk bestående af 4-6 sukkerarter samles k-enheden på lipidbærermolekylet undecaprenylpyrophosphat (Und-P), som giver et stillads til den voksende sukkerkæde (2006hitfield, 2006). Den første sukker i K-enhed, der er ansat i en indre membran (IM)-bundet indledende transferase (Itr), efterfulgt af den sekventielle tilsætning af sukker til den voksende K enhed af specifikke glycosyl transferase (Gtr) enzymer (Figur 1) (Woodward og Naismith, 2016)., Hver k-enhed overføres derefter til den periplasmiske side af IM af the..-translokasen og polymeriseres af. .y, som overfører den voksende polysaccharidkæde fra en Und-p-bærer til den næste indkommende underenhed (Figur 1) (Collins et al., 2007). Efter at CPS-polymeren er syntetiseret, transporteres den til celleoverfladen via en stærkt koordineret proces, der involverer interaktionen mellem tre proteiner;. .a,, .b og. .c, der omfatter eksportmaskineriet (Figur 1)., CPS-syntese repræsenterer en arm af en forgrenet vej, da disse k-enheder også bruges til at dekorere visse overfladeproteiner via O-bundet proteinglycosylering (Lees-Miller et al., 2013). I dette tilfælde overføres enkelt k-enheder til modtagerproteiner af O-oligosaccharyltransferase PglL (Figur 1) (i .ashki.et al., 2012). I A. baumannii, protein glykosylering bidrager til biofilmdannelse ved at styrke oprindelige vedhæftede fil, og modning af biofilm, og patogenicitet, som påvist i en række dyre-infektion modeller (Iwashkiw et al., 2012; Scott et al. , 2014; Harding et al.,, 2015). Biofilm er en væksttilstand, hvor bakteriesamfund er indesluttet i en e .opolysaccharidmatri.og har vist sig at spille en betydelig rolle i A. baumannii Persistens og resistens.
Andre overfladen kulhydrater, der vides at påvirke patogenicitet af A. baumannii omfatter lipooligosaccharide (LOS) og exopolysaccharide poly-β-(1-6)-N-acetylglucosamin (PNAG) (Preston et al., 1996; 1996eber et al., 2016). PNAG danner den sammenhængende” lim ” af biofilm og udgør en betydelig del af biofilm (Choi et al., 2009; Longo et al., 2014)., I modsætning til de fleste Gram-negative bakterier, A. baumannii ikke producerer traditionelle lipopolysaccharide, men i stedet en lignende overflade, glyco-konjugat, LOS, som består af en lipid En kerne, der mangler O-antigen (Kenyon og Hall, 2013; Kenyon et al., 2014b). Tab af LOS produktion i A. baumannii nedsætter stabiliteten af den ydre membran fører til reduceret fitness (Moffatt et al., 2010; Beceiro et al., 2014).
selvom mange kulhydratdele påvirker patogenicitet, kan det hævdes, at CPS er en overvejende virulensfaktor af A. baumannii. Denne gennemgang har til formål at konsolidere det, der er kendt om A., baumannii kapsel, herunder udvalgte strukturer, biosyntese og genorganisation, CPS ‘ rolle i virulens og potentialet for CPS som mål for fremtidig vaccine-og lægemiddeludvikling.
genetisk Organisation af K Loci
efterhånden som de komplette genomer af flere A. baumannii-isolater bliver tilgængelige, bliver den sande mangfoldighed af kapselstrukturer, der findes i denne bakterie, synlig. Til dato er over 100 unikke kapselloci (KL) blevet identificeret i A. baumannii (figur 2A) (Shashkov et al., 2017). Disse regioner spænder typisk fra 20 Til 35 kb i størrelse., Analyse af generne, der dirigerer CPS-syntese i ti komplette A. baumannii-genomer, resulterede oprindeligt i betegnelsen af ni kapseltyper, KL1–KL9, som blev grundlaget for et universelt typeskema for disse loci (Kenyon og Hall, 2013). Denne ordning er efterfølgende udvidet til at imødekomme identifikationen af nye K loci. Den kromosomale placering af K locus, mellem fkpA og lldP gener, er stærkt konserverede mellem A. baumannii stammer og indeholder de gener, der er nødvendige for biosyntese og eksport er specifikke for hver type CPS (Figur 2) (Hu et al.,, 2013; Kenyon og Hall, 2013). En undtagelse fra denne regel er A. baumannii-stammer, der besidder KL19-og KL39-regioner, hvor genet, der koder for polym .y-polymerasen,. .y, findes på en lille genetisk ø andre steder på kromosomet (Kenyon et al., 2016a). Derudover findes de gener, der kræves for nogle af de almindelige sukkerarter, der ses i CPS, andre steder., Alle K loci viser en lignende genetiske konfiguration, en meget variabel klynge af syntese og transferase gener, der er nødvendige for biosyntesen af unikke KL-type komplekse sukkerarter, flankeret på den ene side af den meget velbevarede CPS eksport gener, og på den anden side af et sæt af gener, der koder bevaret simple sukkerarter og eksplosivstoffer (Figur 2A)., Den wzx og wzy gener, der er nødvendige for at gentage-enhed behandlingen er meget varierende mellem K loci (Figur 2A, lys blå) med angivelse af, specificitet for især K-enhed, strukturer, og i almindelighed, rækkefølgen af gtr determinanter, kodning af specifikke glycosyltransferases inden KL regioner, omvendt svarer med henblik på handling.
Den variable region nogle af KL gene clusters, som KL37 og KL14, manglende gener for kompleks sukker syntese, da de kun indeholder de mest simple sukkerarter i deres K-enheder (Arbatsky et al., 2015; Kenyon et al.,, 2015). Også tilføje til deres mangfoldighed, flere KL regioner indeholder overflødige gener, som ikke er nødvendige for syntesen af den endelige k enhed. For eksempel KL8 og KL9 indeholder to itr gener (Kenyon og Hall, 2013) og KL37 har pgt1 phosphoglyceroltransferase, men ingen tilsvarende phosphoglycerol rest i bestemt struktur (Arbatsky et al., 2015). Endvidere Afbryder to insertionssekvenselementer (ISAba26 og ISAba22) i KL93 pgt1-determinanten (figur 2) (Kasimova et al., 2017)., De gener, der kræves til specifik sukkerbiosyntese, vil ikke blive diskuteret, da disse veje tidligere er blevet dækket og ligger uden for denne gennemgang (Hu et al., 2013; Kenyon og Hall, 2013). Derudover andre gener placeret inden for KL kode produkter forudsagt at være involveret i acetylering eller acylering modifikation af specifikke glycaner (figur 2A, pink). Selvom undersøgelse af K loci kan afsløre meget om K-enhedsstrukturer, kemisk analyse og biokemisk test er nødvendig for at bestemme nøjagtige strukturer og identificere specifikke forbindelser mellem gentagne sukkerarter.,
CPS Strukturer
De tidligste undersøgelser af CPS serotypning af A. baumannii blev drevet af behovet for at udvikle en metode til at diskriminere A. baumannii isolater fra andre Acinetobacter arter, som fænotypiske analyser var bebyrdet med tvetydighed og fejlidentifikation (Traub, 1989). Som beskrevet ovenfor er der fænomenal mangfoldighed set i A. baumannii CPS biosyntesegenklynger, som oversætter til mangfoldigheden set i K-enhedsstruktur (Hu et al., 2013; Kenyon og Hall, 2013). Samlet, over 40 forskellige A., baumannii k-enhedsstrukturer er hidtil blevet belyst ved hjælp af NMR-spektroskopi og kemisk analyse. K-enhedsstrukturer adskiller sig i sukkersammensætning. De kan omfatte derivater af almindelige UDP-bundne sukkerarter såsom glucose, galactose og glucuronsyre eller sjældne og atypiske sukkerarter såsom ikke-2-ulosonsyrer. Strukturer varierer i længde og kan bestå af kun to rester, som det ses for K53 type CPS (Shashkov et al., 2018) eller op til fem eller seks monosaccharider, såsom dem, der ses i K37 (figur 2B) (Arbatsky et al., 2015)., Strukturer adskiller sig også i forbindelserne både inden for og mellem K-enheder, hvilket resulterer i produktion af K-enheder, der er lineære eller involverer sidegrener, som det ses i henholdsvis K1 og K93 (figur 2B) (Kenyon et al., 2016a; Kasimova et al., 2017). Forskelle i placeringen af specifikke glycosidbindinger og O-acetyleringsmønstre af forskellige oligosaccharider inden for en struktur bidrager også til K-enhedsdiversitet.,
Variation mellem K-enhedsstrukturer kan være subtil, for eksempel adskiller K12 og K13 kun ved binding af to glycaner, hvilket kræver anvendelse af en alternativ polym .y-polymerase; følgelig er K-loci for begge stammer identiske bortset fra gene .y-genet (figur 2). Alternativt kan variationen være slående, såsom inkorporering af sjældne sukkerarter, herunder pseudamin -, legionaminsyrederivater eller acinetaminsyrederivater som set i henholdsvis K2/6, K49 og K12/13 strukturer (figur 2) (Kenyon et al., 2014a, 2015; Vinogradov et al., 2014; Kasimova et al., 2018)., Interessant nok er acinetaminsyrederivater kun blevet identificeret i A. baumannii og findes intet andet sted i naturen (Kenyon et al ., 2017). Derudover indeholder nogle K-enheder unikke derivater af specifikke glycaner, for eksempel er pseudaminsyren af K93 acetyleret med A (R)-3-hydro .ybutanoylgruppe (Kasimova et al., 2017), mens pseudaminsyren i K2 og K6 ikke er acetyleret (figur 2)., Desuden struktur af K4 er unik, da det kun indeholder aminosugars, D-QuipNAc, og en terminal N-acetyl-D-galactosamine (D-GalpNAcA) filial, som er udjævnet med en pyruvyl gruppe, en sjælden motiv og den første til at blive beskrevet i Acinetobacter (Figur 2A, sort) (Kenyon et al., 2016b). Efterhånden som antallet af K-enhedsstrukturer belyst øges, gør tilliden til at udlede k-enhedsstrukturen fra analyse af biosynteseklyngerne placeret i A. baumannii KL., Selvom det er informativt, er en forståelse af den rolle, CPS spiller i patogenese, vigtig for at anvende denne viden til at forbedre resultaterne af A. baumannii-infektioner.
rolle i virulens, antimikrobiel resistens og persistens
det er uden tvivl, at tilstedeværelsen af CPS er afgørende for A. baumannii-patogenicitet. Ikke alene er det nødvendigt for unddragelse af værtens immunforsvar (Russo et al ., 2010; Geisinger og Isberg, 2015), men det er vigtigt for resistens over for antimikrobielle forbindelser og overlevelse i ugunstige miljøer (Luke et al., 2010; Russo et al. ,, 2010; Geisinger og Isberg, 2015). CPS formidler immununddragelse i mange A. baumannii-stammer ved at begrænse interaktioner mellem immunogene overfladestrukturer af bakterierne og værtsforsvaret (Preston et al., 1996; 1996u et al., 2009; Russo et al. , 2010; Umland et al., 2012; Lees-Miller et al., 2013; Geisinger and Isberg, 2015; Linang-Lin et al., 2017). Afskaffelse af kapsel i flere forskellige A. baumannii stammer har vist reduceret overlevelse i humant serum og ascites væske, og dæmpning i rotte-og mus-infektion modeller (Russo et al., 2010; Umland et al., 2012; Lees-Miller et al.,, 2013; Sanche. – Larrayo. et al., 2017). Desuden op-regulering af kapsel produktion i de almindeligt anvendte A. baumannii stamme ATCC 17978 (K3 CPS-typen), øget serum-resistens og virulens i en mus-infektion model (Geisinger og Isberg, 2015). Desuden kunne nye antimikrobielle behandlinger udvikles til specifikke CPS-typer, for eksempel dem, der indeholder pseudaminsyre, da dets tilstedeværelse er korreleret med forbedret virulens (Hitchen et al., 2010; Kao et al., 2016).
udover beskyttelse mod værtsforsvar, i A., baumannii CPS-produktion øger resistensen over for en række antimikrobielle forbindelser, herunder dem, der anvendes til desinfektion i kliniske omgivelser (Geisinger og Isberg, 2015; Tipton et al., 2015; Chen et al., 2017). Endvidere påvirker væksten af A. baumannii i sub-hæmmende niveauer af antimikrobielle stoffer CPS-produktionen. For eksempel, eksponering for antibiotika chloramphenicol eller erythromycin ført til øget kapsel syntese i ATCC 17978 (Geisinger og Isberg, 2015) og meropenem eksponering, der er valgt for mutationer, der fører til et tab i CPS produktion i de isolere 37662 (Chen et al., 2017)., Undersøgelser udført på en bredere vifte af A. baumannii-stammer er nødvendige for at identificere, om beskyttelsen mod antimikrobielle stoffer, der ydes af CPS, er stammespecifik, kapselspecifik eller universel.
A. baumanniis evne til at fortsætte i det kliniske miljø har utvivlsomt forbedret kolonisering og infektioner hos modtagelige patienter. A. baumannii er i stand til at overleve i flere måneder på hospitalet overflader såsom bed skinner, møbler og medicinsk udstyr, der giver et reservoir, der er ofte kilde til transmission og infektion (Wendt et al., 1997; Gayoso et al., 2013)., Det bemærkelsesværdige udtørring tolerance af A. baumannii menes at være på grund af en “bust” eller boom” – strategi, hvor en vedvarende underbestand af celler overlever på bekostning af døende celler; CPS øger udtørring tolerance ved at skabe en fysisk barriere for at lette vandoptagelse (Roberts, 1996; Webster et al., 2000; Gayoso et al., 2013; Bravo et al., 2016). En direkte rolle for CPS i udtørringsbestandighed blev for nylig demonstreret i A. baumannii-stammen AB5075 (K25 CPS-type). I denne undersøgelse viste den acapsulære variant af AB5075 en 2.,5 gange reduktion i levedygtighed sammenlignet med forældrestammen(Tipton et al ., 2018). Desuden, i to nære slægtninge af A. baumannii, Acinetobacter calcoaceticus og Acinetobacter baylyi, produktion af exopolysaccharide og/eller CPS har vist sig at fremme udtørring overlevelse (Roberson og Firestone, 1992; Ophir og Gutnick, 1994). Ud over at påvirke resistens over for udtørring har CPS været forbundet med andre virulens træk, herunder motilitet (mc .ueary et al., 2012; Huang et al., 2014) og produktion af biofilm (Umland et al ., 2012; Lees-Miller et al.,, 2013), hvilket cementerer sin rolle som en patogen faktor.
de Seneste undersøgelser har knyttet fase-variabel fænotype af A. baumannii AB5075 med ændringer i CPS produktion, som meget virulent uigennemsigtig varianter producere en CPS lag med to gange tykkelsen af deres gennemsigtige kolleger (Chin et al., 2018). Denne overgang fra gennemskinnelig til uigennemsigtig øgede også dramatisk det patogene potentiale af A. baumannii AB5075. Resistens over for almindelige desinfektionsmidler på hospitalet og en undergruppe af aminoglycosidantibiotika blev også øget (Tipton et al., 2015; Chin et al.,, 2018) og uigennemsigtige varianter blev også mere modstandsdygtige over for menneskelige lysozym, den cathelicidin-relaterede antimikrobielle peptid LL37 og hydrogenperoxid i forhold til gennemsigtige kolonier (Chin et al., 2018). Endvidere havde uigennemsigtige isolater en øget tolerance over for udtørrede forhold og udkonkurrerede gennemskinnelige modstykker i en musinfektionsmodel (Chin et al., 2018). Da flere faktorer er involveret i fasevariation, blev der udført yderligere undersøgelser for at bestemme bidraget fra CPS-produktion til den mere virulente uigennemsigtige fænotype., I en efterfølgende publikation demonstrerede forfatterne, at en acapsulær variant var signifikant mere modtagelig for Lyso .ym og desinfektionsmidler sammenlignet med dens uigennemsigtige ab5075 vildtype forælder (Tipton et al., 2018). Interessant nok, var der ingen forskel i resistens over for LL-37 og hydrogenperoxid mellem uigennemsigtig wild-type og acapsular stammer, hvilket tyder på andre faktorer end CPS produktion føre til, at denne fænotype for uigennemsigtig varianter af A. baumannii (Tipton et al., 2018).,
Regulering af CPS-Produktion
Miljømæssige cues, såsom temperatur, osmotisk tryk og ændringer i metabolit og ion tilgængelighed kan påvirke bakteriel CPS produktion (Hagiwara et al., 2003; Lai et al., 2003; Mouslim et al., 2004; 2004illenborg et al., 2011). Det er ikke overraskende, at der kun er identificeret få reguleringsmekanismer for CPS-produktion, da CPS-niveauer ofte reguleres post-translationelt gennem phosphorylering af CPS-eksportmaskiner (Chihitfield og Paiment, 2003; Chiang et al., 2017). I A., baumannii, kun to regulatorer af kapsel produktion har hidtil været identificeret; BfmRS og OmpR-EnvZ, både to-komponent signal transduktion systemer, som spiller flere lovgivningsmæssige roller, der er involveret i konvolut biogenese (Geisinger og Isberg, 2015; Tipton og Snarere 2017; Geisinger et al., 2018). Når det blev udsat for antibiotikumtryk, blev A. baumannii ATCC 17978 CPS-ekspression forøget på en bfmrs-afhængig måde (Geisinger og Isberg, 2015). Fasevariation, og dermed potentielt CPS-produktion, er stærkt reguleret af Ompr-env system-systemet i A., baumannii AB5075, som mutationer i enten ompr eller env.resulterede i en signifikant stigning i uigennemsigtig til gennemskinnelig koblingsfrekvens (Tipton og snarere 2017). Selvom overgang fra gennemskinnelig til uigennemsigtig resulterer i en dobbelt stigning i kapseltykkelse, transkriptomiske analyser har ikke identificeret nogen forskelle i ekspressionsniveauer af KL-gener mellem de to faser.
CPS som mål for udvikling af vacciner og behandlinger mod A. baumannii
antibiotikum (specifikt cabapenem) resistent A., baumannii er klassificeret som en Verdenssundhedsorganisationens Prioritet 1 kritisk organisme til udvikling af nye antimikrobielle stoffer (2017ho, 2017). Selvom der ikke er nogen ikke-antibiotiske behandlinger eller vacciner, der er licenseret til A. baumannii i øjeblikket, er der en øget interesse for deres udvikling, og foreløbige undersøgelser ser lovende ud. Overfladeeksponering og prævalens i patogene stammer af A. baumannii gør CPS til et ideelt mål for både antimikrobielle behandlinger og vacciner (Russo et al., 2013)., Disse inkluderer udvikling af antistofbaserede terapier såsom profylaktiske vacciner, passiv immunisering og fagterapi (Garc .a-.uintanilla et al., 2013).flere undersøgelser har vist effekten af passiv immunisering hos mus ved anvendelse af et CPS-specifikt antistof, som er beskyttende mod bakteriel udfordring med 13-55% af kliniske A. baumannii-isolater (Russo et al., 2013; Yang et al., 2017; Lee et al., 2018)., Derudover, podning med konjugerede vacciner indarbejde CPS glycans knyttet til et protein, der bærer fremkalde bedre immun beskyttelse end renset CPS mod en bred vifte af A. baumannii stammer (Yang et al., 2017).interessen for fagterapi til behandling af bakterielle infektioner er steget i de senere år som reaktion på den aktuelle krise med stigende antimikrobiel resistens. Fagterapi er attraktiv som en potentiel behandlingsmulighed for multidrugsresistente A. baumannii-infektioner., 10%, fire ud af 38, klinisk multidrugsresistent A. baumannii testet (Hernande.-Morales et al., 2018). Selvom værtsområdet for denne fag er begrænset, kan det inkorporeres som en del af en fagcocktail for at maksimere effektiviteten eller målrette specifikke udbrud (Hernande.-Morales et al., 2018). En fag, der selektivt spalter bindingen af A., baumannii CPS ved en pseudaminsyregren kan være værdifuld til fagterapi eller til effektivt at fremstille renset CPS til vaccine-og antistofudvikling (Lee et al., 2018). Fag rettet mod A. baumannii blev for nylig vist at være stabil, når imprægneret i brænde sår plejeprodukter under en række betingelser, herunder i nærvær af antimikrobielle stoffer (Merabishvili et al., 2017). Da størstedelen af undersøgelserne, der undersøger A. baumannii CPS, normalt er begrænset til en bestemt A. baumannii-stamme, vides det ikke, om disse fund oversætter til alle A., baumannii isolerer, eller hvis de er stamme-eller kapseltypespecifikke. At forstå, hvilke roller kapsel spiller i flere stammer, er afgørende for at identificere CPS-typer, der repræsenterer de bedste mål for nye vacciner, eller om udviklingen af antimikrobielle stoffer, der er rettet mod kapselbiosyntesevej, faktisk er mulig.
afsluttende bemærkninger
selvom kapslen repræsenterer et vigtigt virulens træk ved A. baumannii, er der begrænsede data tilgængelige om den rolle, forskellige CPS-typer spiller for at forårsage sygdom., For at udvikle effektive vacciner og terapier, der er rettet mod CPS, skal vi først få en omfattende forståelse for mekanismerne bag dens syntese og ekspression sammen med de fordele, som capsule formidler til værtsbakterierne. Denne forskning skal behandles i sammenhæng med den ekstreme variation af CPS-serotyper, der findes i A. baumannii, for at sikre potentielle interventioner mod stammer, der producerer forskellige CPS-strukturer., Yderligere undersøgelser af CPS er nødvendige for at skabe en platform for udvikling af forebyggende foranstaltninger og behandlinger mod dette stadig mere vedvarende og dødbringende humane patogen.
Forfatterbidrag
JS skrev det første udkast. MB leverede faglige input og kritisk revision af artiklen. FA produceret genom alignments og forudsat kritisk revision af manuskriptet. Alle forfattere godkendte den endelige version.
finansiering
dette arbejde blev støttet af en Flinders Medical Research Foundation-bevilling til mb. FA blev støttet af AJ og Im Naylon og Playford Trust Ph. D., Stipendier. JS blev støttet af en AJ og Im Naylon stipendium.
Erklæring om interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Traub, W. H. (1989). Acinetobacter baumannii serotyping til afgrænsning af udbrud af nosokomial krydsinfektion. J. Clin. Microbiol. 27, 2713–2716.
PubMed Abstract | Google Scholar
Whitfield, C., og Paiment, A. (2003)., Biosyntese og samling af gruppe 1 kapselpolysaccharider i Escherichia coli og beslægtede ekstracellulære polysaccharider i andre bakterier. Carbohydr. Res. 338, 2491-2502. doi: 10.1016/j.carres.2003.08.010
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
DER (2017). Global prioritetsliste over antibiotikaresistente bakterier til vejledning i forskning, opdagelse og udvikling af nye antibiotika. Geneve: Verdenssundhedsorganisationen.
Google Scholar