
introduktion
telomerer er nukleoproteinstrukturer i selve enderne af lineære eukaryote kromosomer. De løser to store end-problemer på samme tid. Den første handler om kromosom endebeskyttelse. Det vurderes, at normale humane celler skal reparere mindst 50 endogene dobbeltstrengede pauser (DSB ‘ er) pr.celle pr. cellecyklus (Vilenchik and Knudson, 2003)., Telomerer adskiller de naturlige kromosomale ender fra skadelige DSB ‘ er og forhindrer deres ektopiske reparation, f.eks. ved ende-til-ende-fusioner af kromosomer (vanSteensel og deLange, 1997). Det andet er slutreplikationsproblemet, der beskæftiger sig med vedligeholdelse af korrekte telomerlængder. Dette blev anerkendt uafhængigt af to forskere (1972atson, 1972; Olovnikov, 1973). Da replikative DNA-afhængige DNA-polymeraser ikke kan fuldføre DNA-syntese i selve enderne af kromosomer, kompensation for replikativ telomersekvens tab skal komme fra en RNA-afhængig DNA-polymerase., Dette enzym, kaldet telomerase, sammen med den første telomere minisatellite sekvens, blev opdaget i cilierede Tetrahymena (Blackburn og Galde, 1978; Greider og Blackburn, 1985). Dette er dog kun et aspekt af vedligeholdelse af telomerlængde. Den epigenetiske regulering af telomerlængde homøostase, herunder samspillet af en lang noncoding telomeric gentag indeholder RNA og exonuclease aktivitet veje, er også blevet grundigt undersøgt på grund af sin terapeutisk potentiale (Wellinger et al., 1996; Polotnianka et al., 1998; Pfeiffer og linger, 2012).,
Telomerase, det en .ym, der er ansvarlig for at tilføje telomere-gentagelsessekvenser til 3′ – enden af telomerer, er et konserveret komplekst en .ym med adskillige komponenter . I princippet er kun to hovedkomponenter essentielle for telomerase-en .ymatisk aktivitet, en katalytisk aktiv proteinkomponent, kaldet telomer revers transkriptase (tert), og en skabelonkomponent, dannet af telomerase RNA-underenheden (tr). Mens tert evolutionært er ret godt bevaret, er TR meget variabel, med længder fra ca. 150 nt (Tetrahymena) til mere end 2.000 nt (svampe fra slægten Neurospora)., Kun en kort region i hele tr-molekylet fungerer som en skabelon til nyligt syntetiseret telomer-DNA (Greider og Blackburn, 1985; .i et al., 2013). Denne region i tr dannes normalt af et komplet telomermotiv efterfulgt af en delvis, sidstnævnte tjener som en udglødningsregion for det eksisterende telomere-DNA. Selvom der i princippet kun er behov for et enkelt ekstra nukleotid (som et delvis motiv), findes der normalt mere end et. For eksempel danner to ekstra nukleotider det udglødende motiv i mus eller fem i mennesker (Blasco et al., 1995; Feng et al., 1995)., I planter er størrelsen af skabelonområdet imidlertid variabel, f.eks. to i Arabidopsis thaliana, syv i Arabis sp. eller seks i Nicotiana (Fajkus et al ., 2019). De øvrige tr-regioner har strukturelle, regulatoriske og proteininteraktive funktioner . Se også en skematisk afbildning af telomerase og dens aktivitetscyklus i Figur 1.

Figur 1 Skematisk fremstilling af telomerase aktivitet cyklus med Arabidopsis-type telomere skabelon. Tert, telomer revers transkriptase; tr, telomerase RNA-underenhed., Figur baseret på Sekhri (2014).
hvor Variable er Telomersekvenser?
Telomersekvenser er normalt korte minisatellitter tandemligt arrangeret, typisk efter formlen (t .ayg.) n.minisatellit-arrangementet stammer fra den måde, hvorpå telomerase syntetiserer DNA ‘ et, kort sagt og for det meste identiske motiver, en efter en., Flere hypoteser, der mener, at en sådan ordning er vigtigt, fordi det fremmer anerkendelse af telomer-specifikke proteiner, homo – og heterodimers og potentiale til at danne G-quadruplexes, der kan stabilisere kromosom-enderne, eller at tjene som underlag for telomer-specifikke proteiner (Spiegel et al., 2020; Tran et al., 2013). Telomersekvenser er godt bevaret gennem evolution, og store grupper af organismer bruger det gruppetypiske telomeremotiv til at opbygge deres telomer-DNA., Et gradvist stigende antal undersøgelser og store screeninger har vist, at alle testede hvirveldyr og mange basale meta .oaner bruger ttaggg (Meyne et al., 1989; Traut et al., 2007) mens Euarthropoda (leddyr), herunder he .apoda (insekter), har TTAGG (Frydrychova et al., 2004; Vitkova et al., 2005). Støt akkumuleres adskillige undtagelser over tid ,F. (A (G)1-8) i Dictyostelium (Emery og andeiner, 1981), ttaggc i Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG in Coleoptera (biller) (Mravinac et al., 2011), TAGGG/TAAGG/TAAGG in Giardia (diplomonader) (U .likova et al.,, 2017), eller TTNNNNAGG i Yarro .ia clade (gær) (Cervenak et al., 2019). Desuden er telomerase-uafhængige systemer, hvor minisatellit-telomersekvensen er gået tabt og erstattet af komplekse gentagelser, for eksempel repræsenteret af Diptera og Chironomidae (gennemgået i (Mason et al., 2016)). For en generel gennemgang af eukaryotisk telomersekvens se (Fajkus et al., 2005; Fulneckova et al., 2013).
telomer sammensætning i planter er endnu mere forskelligartet., Her anvender vi begrebet “anlæg” i bred forstand, også kendt som Archaeplastida eller rige Plantae sensu lato, og bestående af Rhodophyta (rødalger), Glaucophyta, den Chlorophyte alger klasse, og Streptophyte alger klasse (tilsammen kendt som grøn alger), og Embryophyta (jord, planter) (Tusind Plante Transcriptomes Initiativ, 2019). Den typiske telomere plantesekvens er TTTAGGG, også kaldet Arabidopsis-type (eller simpelthen plante-type), da den blev opdaget i Arabidopsis thaliana (Richards og Ausubel, 1988) og nu i mange andre arter på tværs af næsten alle planteordrer., Selvom TTTAGGG stadig er den hyppigste, er der betydelig variation i telomersekvenser i røde og grønne algelinjer. Hvad angår røde alger (Rhodophyta), mangler telomersekvensinformation for det meste eller fragmentarisk, selvom nogle telomerkandidater er blevet opdaget i silico, såsom AATGGGGGG for Cyanidioschy .on merolae (no .aki et al., 2007), TTATT (t) AGG for Galdieria sulphuraria (Fulneckova et al., 2013); TTAGGG er fundet i genomisk læsning af Porphyra umbilicalis (Fulneckova et al., 2013), men flere beviser er nødvendige for at bekræfte deres terminale position på kromosomer., Telomere mangfoldighed i grønne alger afspejler både dynamiske ændringer og dens parafyletiske karakter. Selvom TTTAGGG hersker i Chlorophyta, såsom i slægter Ostreococcus (derelle et al., 2006) og Chlorella (Higashiyama et al ., 1995) er der også fundet mange andre divergerende motiver der, såsom TTAGGG i slægten Dunaliella og Stephanosphaeria (Fulneckova et al., 2012), og TTTTAGGG i Chlamydomonas (Petracek et al., 1990). I basal Streptophyta (Klebsormidiophyceae) progressive ændringer i motiver fra TTTAGGG at TTTTAGGG og TTTTAGG er blevet beskrevet., Tilstedeværelsen af Ttaggg i Rhodophyta og Glaucophyta fører til hypotesen om, at dette er forfædres motiv i planter (Archaeplastida) (Fulneckova et al., 2013).
vedrørende landplanter viste en af de første screeninger, der blev udført, at Arabidopsis-sekvensen var den mest almindelige og for det meste blev bevaret gennem deres fylogeni (Co.et al., 1993; Fuchs et al., 1995), selv om nogle af disse forfattere allerede havde opdaget flere undtagelser i familien Amaryllidaceae (tidligere Alliaceae), hvor Arabidopsis-typen sekvens var fraværende i flere arter., Senere, den første telomere sekvens usædvanligt for jord, planter, hvirveldyr-type TTAGGG, var kendetegnet ved, Aloe og i nogle andre Asparagales (Weiss og Scherthan, 2002; Puizina et al., 2003; Sykorova et al., 2003c). En hypotese om gentagne tab og genfinding af de TTTAGGG og TTAGGG telomere sekvens i Asparagales blev formuleret (Adams et al., 2001). Med postrefinement af orden Asparagaler i APGIII (Angiosperm Fylogeny Group 2009) (Bremer et al.,, 2009), viste det sig, at kun to store evolutionære skifter i telomer sekvens sammensætning opstod (snarere end flere gentagne gevinster og tab), i følgende rækkefølge: den første i familien Iridaceae, hvor et skift fra plante-type TTTAGGG til hvirveldyr-type TTAGGG sket, efterfulgt af familier Xeronemataceae, Asphodelaceae og kernen Asparagales (herunder Amarillidaceae s.l og Asparagaceae s.l.,); og den anden inden for underfamilie Allioideae (tidligere behandlet som en separat familie, Alliaceae), hvor en helt ny telomere sekvens opstået, CTCGGTTATGGG (Fajkus et al., 2016). Uden Asparagales, nye telomer-sekvenser er også blevet påvist i jord-plante-grupper så forskellige som (jeg) Solanaceae, hvor telomere sekvens Busk elegans TTTTTTAGGG blev beskrevet (Sykorova et al., 2003a; Sykorova et al., 2003b; Peska et al., 2008; Peska et al.,, 2015) og (ii) Lentibulariaceae, hvor slægten Genlisea viste en bemærkelsesværdig mangfoldighed med nogle arter er karakteriseret ved Arabidopsis-type telomere gentager, mens andre udstillet blandet rækkefølge varianter TTCAGG og TTTCAGG (Tran et al., 2015).
På trods af alle de konstaterede undtagelser fra telomermotivet er den reelle mangfoldighed i telomersekvenser i landplanter sandsynligvis meget undervurderet. En nylig publikation (Vitales et al .,, 2017), hvor en screening af arealer, anlæg telomer-sekvenser blev udført, fandt, at telomer-sekvenser kun var kendt klart for mindre end 10% af de arter og 40% af de slægter, der er indeholdt i Anlægget rDNA database (www.plantrdnadatabase.com), en ressource, der giver molekylær cytogenetik oplysninger om jord, planter (Garcia et al., 2012). Et resum.af telomersekvensfordeling i planter efter APG IV (Angiosperm Fylogeny Group, 2016) (Byng et al., 2016), såvel som den seneste plantefylogeni (et tusind plante Transkriptomer initiativ, 2019) findes i figur 2.,
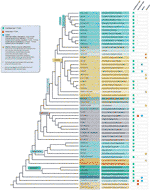
Figur 2 Telomere motiver i Archaeplastida (planter i bred forstand), der er baseret på APG IV (Angiosperm Fylogeni Gruppe 2016) og Tusind Plante Transcriptomes Initiativ (2019). Branch længder ikke udtrykke realtid skalaer., For enkelhed og for at spare plads er visse polyfyletiske “grupper” (kvaliteter) markeret med en stjerne i træet blevet repræsenteret af en enkelt gren; af samme grund er flere mindre ordrer (anført på den blå firkant i venstre overside af figuren) ikke afbildet på træet. Den første tipetiket henviser normalt til planteordrer og i nogle få tilfælde til divisioner, kvaliteter og endda familier; den anden etiket viser repræsentative familier og i nogle få tilfælde repræsentative ordrer eller slægter.
fra screeninger til opdagelse: hvordan telomere motiver kan identificeres?,
beviset for, at en given kandidatsekvens er en reel telomer, inkluderer flere trin, der korrekt erklærer dens lokalisering ved alle kromosomale terminaler og til sidst involvering af telomerase i dens syntese. Molekylær cytogenetik (for det meste ved fluorescens in situ hybridisering, fisk) er blevet vigtig for at visualisere den terminale lokalisering af mærkede prober af kandidatsekvenser ved alle kromosomale terminaler. Men standalone fisk det er ikke nok at bevise den meget terminale position., For eksempel blev AcepSAT356 foreslået i onion som telomerkandidat, baseret på resultater fra FISKEANALYSE (Pich og Schubert, 1998). Ikke desto mindre er dens tilsyneladende terminale placering ved in situ aldrig blevet overbevisende knyttet til telomerfunktion. Faktisk, opdagelsen af Allium minisatellite telomere sekvens CTCGGTTATGGG og telomerase ville betyde, at AcepSAT356 er subterminal (Fajkus et al., 2019)., Positive FISKETELOMERE signaler kan også maskere små ændringer i telomermotiver, såsom enkelt nukleotidpolymorfier, eller falsk-negative resultater kan skyldes, at korte telomerer er under detektionsgrænsen for teknikken.
Der er to yderligere tilgange, der bestemmer terminalpositionen ved større opløsning end fisk; disse er baseret på e .onuclease BAL31-aktivitet., Den første er den klassiske terminale Restriktionsfragment (TRF) – analyse, hvor prøver behandlet med BAL31 viser progressiv forkortelse af terminale fragmenter og et fald i signalintensitet med stigende tid for e .onuclease-behandling. Den efterfølgende analyse af fragment længder udføres ved Southern-blot hybridisering (Fojtova et al., 2015). Den anden er komparativ genom skimming (NGS data) af ikke-fordøjet og BAL31-fordøjet genomisk DNA, parallelt., I BAL31 behandlet datasæt, der er en betydelig underrepræsentation af telomer-sekvenser, derfor terminalen sekvenser er identificeret ved sammenligning med den ubehandlede datasæt, ved hjælp af bioinformatik værktøjer RepeatExplorer eller Tandem Repeats Finder .
den anden vigtige test af en given telomersekvenskandidat i en art er demonstrationen af telomeraseaktivitet. I dette er en nyttig eksperimentel tilgang, der først er udviklet til humane celler, Telomere Repeat Amplification Protocol (TRAP) (Kim et al., 1994), efterfulgt af sekventering af de detekterede produkter (Peska et al.,, 2015; Fajkus et al., 2016), hvilket er lidt mindre følsomt over for falsk positive resultater end fisk. Alle disse metoder, herunder fisk (Fuchs et al., 1995; Shibata og hi .ume, 2011) og andre såsom slot-blot hybridisering (Sykorova et al., 2003c), og fælde (Fulneckova et al ., 2012; Fulneckova et al., 2016), kan bruges til at screene for telomerer på tværs af brede grupper af komplekse organismer, herunder planter., Imidlertid, kun en kombination af passende valgte metoder kan overbevisende føre til en konklusion om telomerfunktionen af en kandidatsekvens, da resultaterne er baseret på en enkelt tilgang kan være vildledende. En mere komplet oversigt over strategier for de novo telomere kandidat rækkefølgen identifikation, herunder den meget første forsøg i Tetrahymena (Greider og Blackburn, 1985) er sammenfattet i en metodologisk artikel, med vægt på NGS tilgang, der anvendes i anlæg med meget store genomer (Peska et al., 2017).
er der homoplasi i Telomersekvenser?,
den forfædres telomersekvens menes at være TTAGGG og er den mest almindeligt forekommende på tværs af livets træ (Fulneckova et al., 2013). Alligevel ser det ud til, at hyppigheden af homoplasi i telomermotivudvikling er relativt høj. For eksempel, kort, enkle motiver som plante-type TTTAGGG har optrådt selvstændigt og flere gange i cryptomonads, oomycete svampe, og alveolates; på samme måde, hvirveldyr-type TTAGGG er opstået sekundært i visse grupper af planter (Asparagales, Rodophyta og Chlorophyta alger) (Sykorova et al., 2003c; Fulneckova et al.,, 2012; Fulneckova et al., 2013; Somanathan og Baysdorfer, 2018). Årsagen til, at nogle telomersekvenser er opstået hyppigere end andre, normalt er mere komplekse sekvenser sandsynligvis relateret til selektionstryk, hvilket ville favorisere nøjagtighed for en bestemt sekvensspecifik DNA-proteininteraktion (Forstemann et al., 2003). Hvis der var en ændring i hvert telomermotiv, ville interferens i den telomeriske nukleoproteinstruktur nødvendigvis føre til genomstabilitet., Dette er grunden til, at telomersekvenser er så evolutionære stabile, omfattende meget få nye og vellykkede sekvenser, et mønster, der er i overensstemmelse med ideen om gentagne tab og fremkomsten af de typiske telomersekvenser, som foreslået for Asparagaler (Adams et al., 2001).
opdagelsen af homoplasi på tværs af telomersekvenser rejser spørgsmålet, Hvad er de molekylære årsager og processer, der finder sted under disse skift? En ændring i telomersekvens, på trods af tilsyneladende triviel i nogle tilfælde (f. eks.,, en ekstra T), kan forårsage alvorlig interferens med genomintegritet på grund af en forstyrret balance i telomere DNA-proteininteraktioner. Det er også uklart, om en ændring i telomersekvens kan have nogen evolutionær fordel; i denne henseende (Tran et al., 2015) foreslog, at udseendet af en “methylerbar” cytosin i en G-rig telomerstreng ville øge muligheden for regulering ved epigenetisk modifikation.
Hvad er de molekylære årsager til ændringer i Telomermotiverne?,
for at forklare ændring af telomersekvens er den første kandidat skabelonunderenheden af telomerase, telomerase RNA (tr). De tidligere identificerede TR fra gær og hvirveldyr tilhører en anden gruppe af afskrifter, hvis tilslutning indslag var, at de blev transskriberet af RNA-polymerase II (Pol II)—i alle, men ciliater; dette bruges til at være den eneste undtagelse fra Pol II udskrifter før offentliggørelsen af den jord, plante TR identifikation . Ved at bruge det relativt lange telomermotiv fra Allium til at kigge efter dets TR inden for den samlede RNA-sekvensdatapool, Fajkus et al., (2019) viste, at et tidligere karakteriseret noncoding RNA involveret i stressreaktionen i A. thaliana, kaldet atr8, var faktisk telomerase RNA-underenheden (.u et al., 2012; Fajkus et al., 2019). Det var et transkript af RNA-polymerase III (Pol III) indeholdende de tilsvarende regulatoriske elementer i dets promotorstruktur. I lang tid forventede forskere, at plante tr ville være så divergerende, at det ville være umuligt at identificere det baseret på en homologi søgning (Cifuentes-Rojas et al., 2011)., Imidlertid, en vis grad af lighed blev med succes brugt til at identificere en fælles TR i flere Allium-arter med sammenlignende eksplosion. Overraskende, sekvens homologi, at tilstedeværelsen af de samme regulerende elementer, og en tilsvarende skabelon regionen førte til identifikation af TRs i Allium, Arabidopsis og mere end 70 andre fjernt beslægtede planter, herunder dem med sig telomere motiver som Genlisea, Busk, og Tulbaghia., Så vidt vi ved, er der stadig ingen data om alger TR, hvilket vil belyse, om Pol III-transkription af tr er et generelt træk for alle planter eller ej. Dette arbejde (Fajkus et al ., 2019), baseret på CRISPR knock-out og andre eksperimenter, viste også, at en tidligere identificeret telomerase RNA kandidat i A. thaliana (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) var ikke en funktionel skabelonunderenhed af telomerase, som det også blev demonstreret kort efter af (de.-Budd et al., 2019)., Forudsat at pol II/Pol III-afhængigheden for tr-transkription er en pålidelig evolutionær markør, vil fremtidig tr-forskning i andre vigtigste eukaryote linjer sandsynligvis åbne ny indsigt i oprindelsen af eukaryoter. Telomerasegener og telomersekvenser er ukendte informationskilder i denne retning, og fundet af en Pol III-afhængig tr-biogenesevej i ciliate og plantelinjer kan repræsentere de første trin i denne retning (Greider og Blackburn, 1989; Fajkus et al., 2019).
Hvordan blev kromosomer lineære?,
et stort flertal af prokaryoter indeholder cirkulære kromosomer, mens lineære kromosomer er reglen i eukaryoter., Der er derfor to mulige scenarier, der enten (i) linearisering blev udført af en primitiv telomerase, forud for andre processer, som førte til den nuværende lineære kromosomale features og funktioner eller (ii) linearisering af en pre-eukaryote cirkulært kromosom blev oprindeligt telomerase uafhængige, men lige før nuværende eukaryoter afveg, en primitiv telomerase begyndte at besætte kromosom-enderne og blev afgørende for den nydannede lineære kromosomer (Nosek, et al., 2006). Villasante et al., (2007) foreslået en evolutionær scenario, hvor brud af slægts-prokaryote cirkulært kromosom aktiveret en gennemførelse mekanisme ved DNA-ender, så dannelsen af telomerer af en rekombination, der er afhængige replikation mekanisme: konsekvenser af denne hypotese førte til den overraskende konklusion, at eukaryote centromeres var afledt af telomerer.
interessant nok er den modsatte proces til linearisering, dvs.dannelse af cirkulære kromosomer (også kaldet ringkromosomer) opstået fra tid til anden under udviklingen af eukaryoter, skønt de er meget ustabile., For eksempel, i tilfælde af Amaranthus tuberculatus, ring kromosomer dukkede op som en stress-induceret svar, der transporterer resistens over for et herbicid (glyphosat); disse ekstra ring kromosomer ikke vise hybridisering med telomere sonder i karyotype analyse (Koo et al., 2018). Det næsten universelle telomerasesystem og undtagelsen af cirkulære kromosomer i eukaryoter tillader os ikke at støtte en hypotese over den anden., Det rekombinationelle maskiner, der anvendes til alternativ forlængelse af telomerer (ALT), en telomerase-uafhængig vej, der er forbundet med visse humane kræftformer (.hang et al., 2019), er allerede til stede i prokaryoter. Derudover er der tegn på kromosomlinearisering, der forekommer uafhængigt i forskellige prokaryote-linjer (Ferdo .s og Barbour, 1989; Nosek et al., 1995; Volff og Altenbuchner, 2000). Derfor synes hypotesen om, at det første lineære eukaryote kromosom (stammende fra en prokaryote-forfader) var telomerase-uafhængig, mere sandsynligt., Der er nogle eksempler, der viser, at telomerase-baseret system er ikke afgørende for telomere vedligeholdelse i alle eukaryoter: retrotransposons i Drosophila telomerer, satellit-gentagelser i Chironomus, et andet insekt (Rubin, 1978; Biessmann og Mason, 2003), og ALT i telomerase-negative menneskelige kræftformer (Hu et al., 2016; 2016hang et al., 2019). Alligevel er nogle af disse systemer måske ikke så forskellige og kan måske have en fælles oprindelse: i Drosophila er telomervedligeholdelsen, der er baseret i retrotransplantation, ikke for forskellig fra den telomerasebaserede mekanisme (Danilevskaya et al.,, 1998), hvilket fører til hypotesen om, at telomerasen selv kan være en tidligere retrotransposon. Men bestemt er telomerase-negative plantearter ikke blevet opdaget til dato, og alle undtagelser, hvor den typiske plante-Type telomer var fraværende, blev senere vist at have forskellige, men stadig telomerase-syntetiserede motiver. Ikke desto mindre er alt-maskinerne til stede i planter parallelt med telomeraseaktiviteten (2007atson og Shippen, 2007; Ruckova et al., 2008). Interessante spørgsmål om telomerasens rolle, telomerer og deres vedligeholdelse i plantetumorer stammer fra det., En attraktiv handler om fraværet af metastase i planter, på trods af tilstedeværelsen af ALT, måske relateret til plantevævsstivhed eller forskellige immunsystemer end hos dyr (Seyfried og Huysentruyt, 2013).
selvom vi får stigende viden om telomerbiologi, er vi stadig ikke i stand til at forklare fremkomsten af telomerase i eukaryoter., Aktuelle beviser understøtter hypotesen om, at fremkomsten af eukaryoter sammen med deres lineære kromosomer, telomerer, og telomerase var relateret til forekomsten af spliceosomal introns archaeask værter (Koonin, 2006; Fajkus et al., 2019). Ligheden mellem tert og andre retroelementer er blevet diskuteret i nogen tid (Pardue et al., 1997). Bemærkelsesværdigt viste en relativt nylig undersøgelse, at tert, som et sandsynligt medlem af afkom gruppe II introner, er sekventielt tæt på Penelope-lignende element retrotransposons (Gladyshev og Arkhipova, 2007)., Men tert er kun en af de to essentielle telomerasekomponenter, og tr er i sin oprindelse endnu mere gåtefuld på grund af dens lave sekvensbeskyttelse på tværs af alle eukaryoter .
konklusion
i begyndelsen af planten genomics æra blev telomersekvensen betragtet som næsten uforanderlig. Den generelle bevarelse af telomerer og telomerasesystemet antydede, at alle planter kan have tttaggg-telomere af plantetype. Identifikation af usædvanlige telomersekvenser i komplekse plantegenomer, i mange tilfælde med kæmpe C-værdier (såsom i Cestrum og Allium sp.,), var umagen værd, da det usædvanligt lange Allium telomere-motiv var ledetråden i at lede efter en ægte TR i landplanter. Den nyligt beskrevet TR i planter og yderligere telomere/telomerase forskning i basal klader af alger kan afsløre værdifuld information om tidlige udvikling, derfor plante telomere forskning kan bidrage væsentligt til en hypotese om fremkomsten af eukaryoter.
Forfatterbidrag
VP og SG har ligeledes bidraget til skrivning, redigering og forberedelse af denne mini-anmeldelse.,
finansiering
interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
et tusind plante Transkriptomer initiativ (2019). Et tusind plante transkriptomer og fylogenomics af grønne planter. Natur 574 (7780), 679-685. doi: 10.1038/s41586-019-1693-2
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar