
introdução
telômeros são estruturas nucleoproteínas nos extremos dos cromossomas eucarióticos lineares. Resolvem dois grandes problemas finais ao mesmo tempo. O primeiro é sobre a proteção final cromossômica. Estima-se que as células humanas normais devem reparar pelo menos 50 fracturas de cadeia dupla endógenas (DSBs) por célula por ciclo celular (Vilenchik e Knudson, 2003)., Os telómeros distinguem as extremidades cromossómicas naturais dos DSBs nocivos e impedem a sua reparação ectópica, por exemplo, através de fusões finais de cromossomas (vanSteensel e deLange, 1997). O segundo é o problema de replicação final que lida com a manutenção de comprimentos telômeros adequados. Isto foi reconhecido independentemente por dois pesquisadores (Watson, 1972; Olovnikov, 1973). Uma vez que as ADN-polimerases replicativas dependentes do ADN não podem completar a síntese de ADN nas extremidades dos cromossomas, a compensação pela perda da sequência de telómeros replicativos deve provir de uma ADN-polimerase dependente do ARN., Esta enzima, chamada telomerase, juntamente com a primeira sequência telomere minissatellite, foi descoberta na Tetrahymena ciliada (Blackburn e Gall, 1978; Greider e Blackburn, 1985). No entanto, este é apenas um aspecto da manutenção de comprimento telômero. A regulação epigenética da homeostase do comprimento do telômero, incluindo a interacção de longas vias de repetição telomérica não codificante contendo RNA e vias de actividade das exonucleases, também foi extensivamente estudada devido ao seu potencial terapêutico (Wellinger et al., 1996; Polotnianka et al., 1998; Pfeiffer e Lingner, 2012).,a Telomerase, a enzima encarregada de adicionar sequências de repetição de telômero à extremidade 3′ de telómeros, é uma enzima complexa conservada com numerosos componentes . Em princípio, apenas dois componentes principais são essenciais para a actividade enzimática da telomerase, um componente proteico cataliticamente activo, chamado telomere reverse transcriptase (terc), e um componente template, formado pela subunidade de RNA telomerase (TR). Enquanto TERT é evolutivamente bastante bem conservado, TR é muito variável, com comprimentos que variam de ca. 150 nt (Tetrahymena) a mais de 2.000 nt (fungos do género Neurospora)., Apenas uma pequena região em toda a molécula TR serve como um modelo para o recém-sintetizado DNA telômero (Greider e Blackburn, 1985; Qi et al., 2013). Esta região em TR é normalmente formada por um motivo telômero completo seguido por um motivo parcial, o último servindo como uma região de recozimento para o DNA telômero existente. Embora, em princípio, apenas um nucleótido extra é necessário (como um motivo parcial), geralmente mais de um é encontrado. Por exemplo, dois nucleótidos extra formam o motivo de recozimento em ratos ou cinco em humanos (Blasco et al., 1995; Feng et al., 1995)., Em plantas, no entanto, o tamanho da região modelo é variável, por exemplo, dois em Arabidopsis thaliana, sete em Arabis sp. ou seis em Nicotiana (Fajkus et al., 2019). As outras regiões TR têm funções estruturais, regulamentares e interactivas de proteínas . Ver também uma representação esquemática da telomerase e do seu ciclo de actividade na Figura 1.

a Figura 1 representação Esquemática do ciclo de atividade da telomerase com Arabidopsis tipo de telômero modelo. Terc, Telomere Reverse Transcriptase; TR, telomerase RNA subunit., Figura baseada em sekhri (2014).
quão variáveis são as sequências de Telomere?
sequências de Telomere são geralmente Mini-satélites curtos dispostos em tandemly, tipicamente seguindo a fórmula (TxAyGz) n. o arranjo minissatélita origina-se da forma em que telomerase sintetiza o DNA, em suma, e na maioria dos motivos idênticos, um por um. , Várias hipóteses consideram que tal arranjo é importante porque promove o reconhecimento de proteínas específicas de telômero por homo-e heterodímeros e para o potencial de formar quadruplexos G que podem estabilizar as extremidades dos cromossomos ou servir como substratos para proteínas específicas de telômero (Spiegel et al., 2020; Tran et al., 2013). As sequências de Telomere são bem conservadas através da evolução, e grandes grupos de organismos usam o motivo telomere típico de grupo para construir seu DNA telomere., Um número cada vez maior de estudos e de grandes testes mostraram que todos os vertebrados testados e muitos metazoanos basais usam TTAGG (Meyne et al., 1989; Traut et al., 2007), enquanto Euarthropoda (artrópodes), incluindo Hexapoda (insetos), tem TTAGG (Frydrychova et al., 2004; Vitkova et al., 2005). Gradualmente, as numerosas exceções estão se acumulando ao longo do tempo, por exemplo, (A(G)1-8) Dictyostelium (Emery e Weiner, 1981), TTAGGC em Ascaris lumbricoides (Nematoda) (Muller et al., 1991), TCAGG in Coleoptera (beetles) (Mravinac et al., 2011), Tagg/TAAGG/TAAGG in Giardia (diplomonads) (Uzlikova et al.,, 2017), ou TTNNNNAGG in Yarrowia clade (leveduras) (Cervenak et al., 2019). Além disso, sistemas independentes de telomerase, em que a sequência telomere minissatellite foi perdida e substituída por repetições complexas, são representados, por exemplo, por Diptera e Chironomidae (revisado em (Mason et al., 2016)). Para uma revisão geral sobre a sequência telomérica eucariótica ver (Fajkus et al., 2005; Fulneckova et al., 2013).a composição Telomérica nas plantas é ainda mais diversificada., Aqui usamos o termo “plantas” em um sentido amplo, também conhecido como Archaeplastida ou reino Plantae-graduação lato sensu, englobando Rhodophyta (algas vermelhas), Glaucophyta, o Chlorophyte algas grau e o Streptophyte algas grau (completamente conhecidas como algas verdes), e Embryophyta (terra, plantas) (Mil Planta Transcriptomes Iniciativa, 2019). A sequência típica de plantas telômeras é TTTAGG, também chamado de tipo Arabidopsis (ou simplesmente tipo de planta) desde que foi descoberto em Arabidopsis thaliana (Richards e Ausubel, 1988) e agora em muitas outras espécies em quase todas as ordens de plantas., Embora o TTTAGG seja ainda o mais frequente, existe uma variabilidade significativa nas sequências de telômeros nas linhagens de algas vermelhas e verdes. Quanto às algas vermelhas (Rhodophyta), a informação da sequência telomere está em falta ou fragmentada, embora alguns candidatos de telomere tenham sido descobertos em slico, como AATGGGG para Cyanidioschyzon merolae (Nozaki et al., 2007), TTATT(T) AGGG for Galdieria sulphuraria (Fulneckova et al., 2013); TTAGG foi encontrado em leituras genômicas de Porphyra umbilicalis (Fulneckova et al., 2013), mas mais evidências são necessárias para confirmar a sua posição terminal nos cromossomas., A diversidade de telómeros em algas verdes reflecte tanto as mudanças dinâmicas como o seu carácter parafilético. Embora TTTAGG prevaleça na Clorofita, como no gênero Ostreococcus (Derelle et al., 2006) e chlorela (Higashiyama et al., 1995), muitos outros motivos divergentes foram detectados lá também, como TTAGG no gênero Dunaliella e Stephanosphaeria (Fulneckova et al., 2012), e TTTTAGG in Chlamydomonas (Petracek et al., 1990). Em streptophyta basal (Klebsormidiophyceae) foram descritas alterações progressivas nos motivos para a mudança de TTTAGG para TTTAGG e TTTAGG., A presença de TTAGG em Rhodophyta e Glaucophyta leva à hipótese de que este é o motivo ancestral das plantas (Archaeplastida) (Fulneckova et al., 2013).
no que diz respeito às plantas terrestres, uma das primeiras exibições realizadas mostrou que a sequência Tipo Arabidopsis era a mais comum e foi conservada principalmente através de sua filogenia (Cox et al., 1993; Fuchs et al., 1995), embora alguns destes autores já tivessem detectado várias exceções na família Amaryllidaceae (antiga Alliaceae), na qual a sequência do tipo Arabidopsis estava ausente em várias espécies., Mais tarde, a primeira sequência telomere incomum para plantas terrestres, o tipo vertebrado TTAGG, foi caracterizada em Aloe e em alguns outros Asparagales (Weiss e Scherthan, 2002; Puizina et al., 2003; Sykorova et al., 2003c). Uma hipótese sobre perdas repetidas e recuperações da sequência telomere de TTTAGG e TTAGG em Asparagales foi formulada (Adams et al., 2001). With the postrefinement of order Asparagales in the APGIII (Angiosperm Phylogeny Group 2009) (Bremer et al.,, 2009), foi mostrado que apenas dois grandes evolutiva opções em telômero sequência de composição ocorreu (em vez de várias repetidas perdas e ganhos), na seguinte ordem: o primeiro na família Iridaceae, em que a transição de uma planta-tipo TTTAGGG para os vertebrados-tipo TTAGGG aconteceu, seguido por famílias Xeronemataceae, Asphodelaceae e o núcleo Asparagales (incluindo Amarillidaceae s.l e Asparagaceae s.l.,); e o segundo dentro da subfamília Allioideae (anteriormente tratada como uma família separada, Alliaceae) em que uma sequência telomere completamente nova emergiu, CTCGGTTATGG (Fajkus et al., 2016). Fora de Asparagales, novas sequências de telômero também foram detectadas em grupos de plantas terrestres tão díspares como (I) Solanaceae, em que a sequência telomere de Cestrum elegans TTTTAGG foi descrita (Sykorova et al., 2003a; Sykorova et al., 2003b; Peska et al., 2008; Peska et al.,, 2015) and (ii) Lentibulariaceae, where genlisea showed a remarkable diversity with some species charactered by the Arabidopsis-type telomere repeats while others exhibited intermingled sequence variants TTCAGG and TTTCAGG (Tran et al., 2015).apesar de todas as exceções teloméricas detectadas, a verdadeira diversidade das sequências teloméricas nas plantas terrestres está provavelmente muito subestimada. Uma publicação recente (Vitales et al.,, 2017), em que uma proteção de terra a planta dos telómeros sequências foi realizada, constatou que o telômero sequências só foram conhecidas claramente para menos de 10% das espécies e 40% dos gêneros contidos na Planta de rDNA (banco de dados www.plantrdnadatabase.com), um recurso de prestação de citogenética molecular informações sobre as plantas terrestres (Garcia et al., 2012). A summary of telomere sequence distribution in plants, following APG IV (the Angiosperm Phylogeny Group, 2016) (Byng et al., 2016), bem como a mais recente filogenia vegetal (iniciativa Mil transcriptomas vegetais, 2019) é encontrada na Figura 2.,
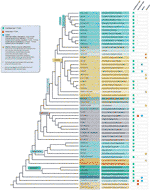
Figure 2 Telomere motifs in Archaeplastida (plants in the broad sense), based on the APG IV (the Angiosperm Phylogeny Group 2016) and on the one Thousand Plant Transcriptomes Initiative (2019). Os comprimentos dos ramos não exprimem escalas em tempo real., Para a simplicidade e para economizar espaço, certas polyphyletic “grupos” (graus) marcados com um asterisco na árvore de ter sido representado por um único ramo; pela mesma razão, várias ordens menores (listados no quadrado azul no lado superior esquerdo da figura) não são representadas na árvore. A primeira etiqueta de ponta refere-se geralmente às encomendas de plantas e, em alguns casos, a divisões, graus e até mesmo famílias; a segunda etiqueta mostra famílias representativas e, em alguns casos, ordens representativas ou géneros.
de exibições à descoberta: como motivos Teloméricos podem ser identificados?,
a evidência de que uma dada sequência candidata é uma telomérica real inclui várias etapas que declaram corretamente sua localização em todas as terminações cromossômicas, e eventualmente o envolvimento da telomerase em sua síntese. A citogenética Molecular (principalmente por fluorescência in situ hibridização, FISH) tornou-se importante para visualizar a localização terminal de sondas rotuladas de sequências candidatas em todos os terminos cromossômicos. No entanto, os peixes independentes não são suficientes para provar a posição muito terminal., Por exemplo, AcepSAT356 foi proposto em onion como o candidato telomere, com base nos resultados da análise de peixes (Pich e Schubert, 1998). No entanto, a sua aparente localização terminal Por in situ nunca foi convincentemente ligada à função telomere. Na verdade, a descoberta da sequência telomere CTCGGTTATGGG e telomerase do Allium minissatellite significaria que AcepSAT356 é um subconjunto (Fajkus et al., 2019)., Sinais telômeros de peixes positivos também podem mascarar pequenas alterações em motivos telômeros, tais como polimorfismos de nucleótidos únicos, ou resultados falsos negativos podem resultar de telômeres curtos estarem abaixo do limite de detecção da técnica.
Existem duas abordagens adicionais que determinam a posição terminal em maior resolução do que os peixes; estas são baseadas na actividade da BAL31 da exonuclease., A primeira é a análise clássica do fragmento de restrição Terminal (TRF), na qual amostras tratadas por BAL31 mostram redução progressiva de fragmentos terminais e diminuição da intensidade do sinal com o aumento do tempo de tratamento da exonuclease. A análise subsequente dos comprimentos dos fragmentos é realizada pela hibridização Southern-blot (Fojtova et al., 2015). O segundo é o estudo comparativo do genoma skimming (dados NGS) do DNA genômico não digerido e BAL31 digerido, em paralelo., No conjunto de dados Tratado de BAL31, existe uma sub-representação significativa das sequências de telômero, portanto as sequências terminais são identificadas por comparação com o conjunto de dados não tratados, usando ferramentas bioinformáticas Repetexplorer ou repetições Tandem Finder .
o outro teste importante de uma dada sequência telomere candidata numa espécie é a demonstração da actividade da telomerase. Neste, uma abordagem experimental útil, desenvolvida primeiro para as células humanas, é o protocolo de amplificação repetitiva Telomere (TRAP) (Kim et al., 1994), seguida pela sequenciação dos produtos detectados (Peska et al.,, 2015; Fajkus et al., 2016), que é um pouco menos sensível aos resultados falsos positivos do que o peixe. Todos estes métodos, incluindo o peixe (Fuchs et al., 1995; Shibata and Hizume, 2011) and others such as slot-blot hybridization (Sykorova et al., 2003c), and TRAP (Fulneckova et al., 2012; Fulneckova et al., 2016), pode ser usado para rastrear telômeros através de grandes grupos de organismos complexos, incluindo plantas., No entanto, apenas uma combinação de métodos adequadamente escolhidos pode levar convincentemente a uma conclusão sobre a função telomere de uma sequência candidata, uma vez que os resultados baseados numa única abordagem podem ser enganadores. Uma visão mais completa das estratégias de novo telômero candidato seqüência de identificação, incluindo a primeira tentativa em Tetrahymena (Greider e Blackburn, 1985) são resumidas em um artigo metodológico, com ênfase na NGS abordagem utilizada em plantas com extremamente grandes genomas (Peska et al., 2017).há Homoplasy nas sequências Teloméricas?,
acredita-se que a sequência telomérica ancestral seja TTAGG e é a mais comumente encontrada através da árvore da vida (Fulneckova et al., 2013). No entanto, parece claro que a frequência de homoplasy na evolução telomere motif é relativamente alta. Por exemplo, curto, simples motivos como a planta-tipo TTTAGGG têm aparecido de forma independente e repetidamente em cryptomonads, oomycete fungos, e alveolates; da mesma forma, o vertebrados-tipo TTAGGG surgiu, secundariamente, em certos grupos de plantas (Asparagales, Rodophyta e algas Chlorophyta) (Sykorova et al., 2003c; Fulneckova et al.,, 2012; Fulneckova et al., 2013; Somanathan and Baysdorfer, 2018). A razão pela qual alguns dos telómeros sequências têm surgido com mais frequência do que outras, geralmente mais complexas sequências provavelmente está relacionado a pressões seletivas, que favoreçam a precisão de uma determinada sequência específica de DNA-proteína de interação (Forstemann et al., 2003). Se houvesse uma mudança em cada motivo telômero, a interferência na estrutura da nucleoproteína telomérica levaria necessariamente à instabilidade do genoma., Esta é a razão pela qual as sequências de telômero são tão evolutivas estáveis, compreendendo muito poucas sequências novas e bem sucedidas, um padrão consistente com a ideia de perdas repetidas e o surgimento das sequências telômeras típicas, como proposto para Asparagales (Adams et al., 2001).
a descoberta de homoplasy através de sequências telômeras levanta a questão, Quais são as causas e processos moleculares que ocorrem durante estas mudanças? Uma mudança na sequência de telômero, apesar de parecer trivial em alguns casos (e.g.,, um T extra), pode causar interferência grave na integridade do genoma, devido a um equilíbrio perturbado nas interações ADN-proteína telomere. Também não é claro se uma mudança na sequência telômero pode ter alguma vantagem evolutiva; a este respeito, (Tran et al., 2015) sugeriu que o aparecimento de uma citosina “metilável” em uma cadeia telomérica Rica Em G aumentaria a possibilidade de regulação por modificação epigenética.
quais são as razões moleculares para as alterações nos motivos de Telomere?,
para explicar a alteração da sequência telomere, o primeiro candidato é a subunidade modelo da telomerase, telomerase RNA (TR). O TR previamente identificado a partir de leveduras e vertebrados pertence a um grupo diferente de transcrições, cuja característica de conexão era que eles foram transcritos por RNA polimerase II (Pol II)—em todos, exceto ciliates; esta costumava ser a única exceção de transcrições Pol II antes da publicação da identificação da planta terrestre TR . Usando o motivo telômero relativamente longo do Allium para procurar o seu TR dentro do conjunto de dados de sequência de RNA total, Fajkus et al., (2019) showed that a previously characterized noncoding RNA in the stress reaction in A. thaliana, called AtR8, was indeed the telomerase RNA subunit (Wu et al., 2012; Fajkus et al., 2019). Foi uma transcrição da RNA polimerase III (Pol III) contendo os elementos regulamentares correspondentes na sua estrutura Promotora. Durante muito tempo, pesquisadores esperavam que a planta TR fosse tão divergente que seria impossível identificá-la com base em uma pesquisa de homologia (Cifuentes-Rojas et al., 2011)., No entanto, um certo grau de semelhança foi usado com sucesso para identificar um TR comum em várias espécies de Allium com explosão comparativa. Surpreendentemente, a seqüência de homologia, a presença dos mesmos elementos de regulamentação, e um modelo correspondente região levou à identificação de TRs em Allium, Arabidopsis e mais de 70 outros aparentados plantas, incluindo aqueles com divergiram dos telómeros motivos como Genlisea, Cestrum, e Tulbaghia., Tanto quanto sabemos, ainda não há dados sobre qualquer alga TR, que esclareçam se a transcrição de RS por Pol III é uma característica geral para todas as plantas ou não. Este trabalho (Fajkus et al., 2019), baseado em CRISPR knock-out e outros experimentos, também mostrou que um candidato a RNA telomerase previamente identificado em A. thaliana (Cifuentes-Rojas et al., 2011; Beilstein et al., 2012) não foi uma subunidade de modelo funcional da telomerase, como também foi demonstrado pouco depois por (Dew-Budd et al., 2019)., Assumindo que a dependência Pol II/Pol III da transcrição TR é um marcador evolucionário confiável, a futura pesquisa TR em outras linhagens eucarióticas principais provavelmente abrirá novas perspectivas sobre a origem dos eucariontes. Os genes da Telomerase e as sequências teloméricas são fontes não reconhecidas de informação nesta direcção, e a descoberta de uma via de biogénese TR dependente de Pol III em linhagens ciliadas e vegetais pode representar os primeiros passos nesta direcção (Greider e Blackburn, 1989; Fajkus et al., 2019).como os cromossomas se tornaram Lineares?,
uma grande maioria dos procariontes contém cromossomas circulares, enquanto os cromossomas lineares são a regra nos eucariontes., Portanto, há dois cenários possíveis em que (i) linearização foi realizada por uma primitiva da telomerase, anterior outros processos que levaram à atual linear cromossômicas características e funções ou (ii) de linearização de um pré-eucarióticas circular cromossoma foi inicialmente telomerase independente, mas antes de a atual eucariotas divergiu, um primitivo telomerase começaram a ocupar as pontas dos cromossomas e se tornou essencial para os recém-formados linear cromossomos (Nosek et al., 2006). Villasante et al., (2007) propuseram um cenário evolucionista, em que a quebra do ancestral procariontes circular cromossomo activado um mecanismo de transposição no DNA extremidades, o que permite a formação de telômeros por uma recombinação dependentes do mecanismo de replicação: conseqüências desta hipótese levou à surpreendente conclusão de que eucarióticas centromeres foram obtidas a partir de telômeros.curiosamente, o processo oposto à linearização, ou seja, a formação de cromossomas circulares (também chamados cromossomas anelares) emergiu de tempos em tempos durante a evolução dos eucariotas, embora sendo altamente instável., Por exemplo, no caso do amaranto tuberculatus, os cromossomas do anel apareceram como uma resposta induzida por stress, Carregando resistência contra um herbicida (glifosato); estes cromossomas do anel extra não mostraram hibridização com sondas de telomere na análise do cariótipo (Koo et al., 2018). O sistema quase universal de telomerase e a excepcionalidade dos cromossomas circulares nos eucariontes não nos permitem apoiar uma hipótese sobre a outra., No entanto, a máquina recombinacional utilizada no prolongamento alternativo dos telómeros (ALT), uma via independente da telomerase, associada a certos cancros humanos (Zhang et al., 2019), já está presente em procariontes. Além disso, há evidências de linearização cromossômica ocorrendo independentemente em linhagens procariotas distintas (Ferdows e Barbour, 1989; Nosek et al., 1995; Volff e Altenbuchner, 2000). Portanto, a hipótese de que o primeiro cromossoma eucariótico linear (originário de um ancestral procariota) era independente da telomerase parece mais provável., Há alguns exemplos que mostram que a telomerase baseado no sistema não é essencial para a manutenção dos telómeros em todos os eucariontes: retrotransposons em Drosophila telômeros, satélite se repete em Chironomus, outro inseto (Rubin, 1978; Biessmann e Mason, 2003), e ALT em telomerase-negativo cânceres humanos (Hu et al., 2016; Zhang et al., 2019). No entanto, alguns destes sistemas podem não ser tão diferentes, e talvez possam ter uma origem comum: em Drosophila, a manutenção telomere, baseada na retrotransposição, não é muito distinta do mecanismo baseado na telomerase (Danilevskaya et al.,, 1998), levando à hipótese de que a telomerase em si pode ser um antigo retrotransposon. Mas certamente, espécies vegetais telomerase-negativas não foram descobertas até à data e todas as exceções, nas quais o típico telomere tipo planta estava ausente, foram mais tarde mostradas como tendo motivos diferentes, mas ainda sintetizados telomerase. No entanto, a maquinaria ALT está presente em instalações paralelas à actividade da telomerase (Watson e Shippen, 2007; Ruckova et al., 2008). Questões interessantes sobre o papel da telomerase, telômeres e sua manutenção em tumores de plantas surgem a partir disso., Um atrativo é sobre a ausência de metástases nas plantas, apesar da presença de ALT, talvez relacionada com a rigidez dos tecidos vegetais ou diferentes sistemas imunológicos do que em animais (Seyfried e Huysentruyt, 2013).apesar de estarmos ganhando cada vez mais conhecimento da biologia telomérica, ainda não conseguimos explicar o surgimento da telomerase em eucariontes., Evidências atuais sustentam a hipótese de que o aparecimento de eucariontes juntamente com seus cromossomos lineares, telômeres e telomerase estava relacionado com o aparecimento de intrões spliceossômicos em hospedeiros archaeal (Koonin, 2006; Fajkus et al., 2019). A semelhança entre TERT e outros retroelements tem sido discutido por algum tempo (Pardue et al., 1997). Notavelmente, um estudo relativamente recente mostrou que TERT, como um provável membro dos introns do grupo II da progênie, é sequencialmente próximo de retrotransposons Elemento Tipo Penélope (Gladyshev e Arkhipova, 2007)., Mas TERT é apenas um dos dois componentes essenciais da telomerase, e TR é, em sua origem, ainda mais enigmático devido à sua conservação de baixa sequência em todos os eucariontes .
conclusão
no início da era genómica vegetal, a sequência telomérica foi considerada quase imutável. A conservação geral dos telómeros e do sistema telomerase sugeriram que todas as plantas podem ter o tipo telomere da planta TTTAGG. The identification of unusual telomere sequences in complex plant genomes, in many cases with giant C-values (such as in Cestrum and Allium sp.,), valeu a pena o esforço, uma vez que o motivo excepcionalmente longo de Allium telomere foi a pista na procura de um TR genuíno em plantas terrestres. O TR recentemente descrito em plantas e outras pesquisas telomere/telomerase em clados basais de algas podem revelar informações valiosas sobre a evolução precoce, portanto, a pesquisa telomérica de plantas pode contribuir significativamente para hipóteses sobre o surgimento de eucariontes.
contribuições dos autores
VP e SG contribuíram igualmente para a escrita, edição e preparação desta mini-revisão.,
financiamento
conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
agradecimentos
1000 Plant Transcriptomes Initiative (2019). Mil transcriptomas vegetais e a filogenómica de plantas verdes. Nature 574 (7780), 679-685. doi: 10.1038 / s41586-019-1693-2
PubMed Abstract | CrossRef Full Text | Google Scholar